Biology:Notoungulata
| Notoungulata | |
|---|---|

| |
| Skeleton of Toxodon (Toxodontidae) | |

| |
| Skeleton of Prosotherium (Hegetotheriidae) | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Clade: | Panperissodactyla |
| Order: | †Notoungulata Roth 1903 |
| Suborders and families | |
|
See text | |
Notoungulata is an extinct order of mammalian ungulates that inhabited South America from the early Paleocene to the Holocene, living from approximately 61 million to 11,000 years ago.[1] Notoungulates were morphologically diverse, with forms resembling animals as disparate as rabbits and rhinoceroses. Notoungulata are the largest group of South American native ungulates, with over 150 genera in 14 families having been described, divided into two major subgroupings, Typotheria and Toxodontia. Notoungulates first diversified during the Eocene. Their diversity declined from the late Neogene onwards, with only the large toxodontids persisting until the end of the Pleistocene (with Mixotoxodon expanding into Central America and southern North America), perishing as part of the Quaternary extinction event among with most other large mammals in the Americas. Collagen analysis suggests that notoungulates are closely related to litopterns, another group of South American ungulates, and their closest living relatives being perissodactyls (odd-toed ungulates), including rhinoceroses, tapirs and equines as part of the clade Panperissodactyla. However their relationships to other South American ungulates are uncertain. Several groups of notoungulates separately evolved ever-growing cheek teeth.
Taxonomy
Notoungulata is divided into two major suborders, Typotheria and Toxodontia, alongside some basal groups (Notostylopidae and Henricosborniidae) which are potentially paraphyletic.[2]
This order is proposed to be united with other South American native ungulates in the super-order Meridiungulata. The notoungulate and litoptern native ungulates of South America have been shown by studies of collagen and mitochondrial DNA sequences to be a sister group to the perissodactyls, making them true ungulates.[3][4][5] The estimated divergence date is 66 million years ago.[5] This conflicts with the results of some morphological analyses which posited them as afrotherians. It is in line with some more recent morphological analyses which suggested they were basal euungulates. Panperissodactyla has been proposed as the name of an unranked clade to include perissodactyls and their extinct South American ungulate relatives.[3]
Cifelli has argued that Notioprogonia is paraphyletic, as it would include the ancestors of the remaining suborders. Similarly, Cifelli indicated that Typotheria would be paraphyletic if it excluded Hegetotheria and he advocated inclusion of Archaeohyracidae and Hegetotheriidae in Typotheria.[6]
Notoungulata were for many years taken to include the order Arctostylopida, whose fossils are found mainly in China. Recent studies, however, have concluded that Arctostylopida are more properly classified as gliriforms, and that the notoungulates were therefore never found outside South and Central America.[7]
Based on an analysis of 133 morphological characters in 50 notoungulate genera, Billet in 2011 concluded that Homalodotheriidae, Leontiniidae, Toxodontidae, Interatheriidae, Mesotheriidae, and Hegetotheriidae are the only monophyletic families of notoungulates. Some studies have suggested that Pyrotheria, often ranked as an independent order, should also be included within Notoungulata.[8]
Phylogeny
| Notoungulata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ecology
Notoungulates varied widely in body size, with early diverging notoungulates like Simpsonotus, and some hegetotheriid and interatheriid typotherians having a body mass of approximately 1–2 kilograms (2.2–4.4 lb), while the toxodontid Toxodon is suggested to have had a body mass exceeding 1,000 kilograms (2,200 lb). Typotheres generally occupied small-medium body size niches, while toxodontians were generally medium-large sized animals.[2] The families Interatheriidae, Hegetotheriidae, Mesotheriidae and Toxodontidae separately evolved high crowned (hypsodont) ever-growing cheek teeth,[9] with high crowned species constiuting the majority of notoungulates from the Late Oligocene onward.[2] This adaptation was historically suggested to be the result of a diet increasingly incorporating grass, but this has been questioned, and other authors suggesting that it may have been due to the increasing intake of abrasive particles from volcanic sources.[2][9] Many typotheres have bodyforms convergent on rodents, hyraxes and rabbits,[10] with some rabbit-like hegetotheriids suggested to have developed a rabbit-like bounding locomotion.[11] The basal notungulate Notostylops and the mesotheriids are suggested to have engaged in digging, with mesotheriids suggested to have had an ecology similar to wombats.[2] Toxodontids have sometimes been compared to rhinoceroses and hippopotamuses in overall bodyform and tooth morphology.[10] The Miocene toxodontian Homalodotherium had claws on its forelimbs and is thought to have had an ecology similar to the extinct chalicotheres.[2] Like perissodactyls, notoungulates were likely primitively hindgut fermenters,[12] but it has also been proposed that some of them may have had fermentation more similar to ruminants based on their skeletal anatomy, though this is uncertain.[10]

Restoration of Thomashuxleya, an early toxodontian notoungulate from the Eocene (Lutetian) of Argentina

Skull of Hegetotherium (Typotheria, Hegetotheriidae)
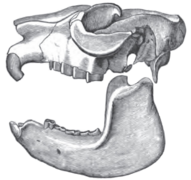
Skull of Mesotherium (Typotheria, Mesotheriidae)

Skeleton of Miocochilius (Typotheria, Interatheriidae)

Skeleton of Nesodon (Toxodontia, Toxodontidae)
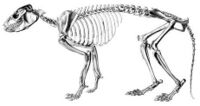
Skeleton of Protypotherium (Typotheria, Interatheriidae)

Skeleton of Pachyrukhos (Typotheria, Hegetotheriidae)

Skeleton of Homalodotherium (Toxodontia, Homalodotheriidae)

Rhynchippus (Toxodontia, Notohippidae)

Scarrittia (Toxodontia, Leontiniidae)










Evolutionary history
The oldest notoungulates appeared during the Paleocene,[1] probably originating from "condylarth" ancestors that had migrated from North America. Notoungulates and other South American native ungulates reached their apex of diversity during the Eocene and Oligocene. Notoungulate species diversity was stable during the Miocene, though 45% of the family diversity of the group became extinct during the interval, including Homalodotheriidae, Leontiniidae, and Interatheriidae. The diversity of the group declined during the Pliocene and Pleistocene, which is coeval in time with the Great American Interchange, which allowed ungulates and other mammals from North America to enter South America. This decline has historically been attributed to competition with the new North American arrivals, though earlier views had probably overstated the importance of this,[2] with climatic change also likely being an important factor.[13] As part of the Great American interchange, the toxodontid Mixotoxodon migrated into Central and North America, with its furthest northern record being in Texas.[14] The last hegetotheriids are known from the Early Pleistocene (with a supposed Middle Pleistocene record being considered questionable).[13] The youngest known member of Typotheria, the mesotheriid Mesotherium, has its last records in the late Middle Pleistocene, around 220,000 years ago.[15] The last notoungulates, the toxodontids Toxodon, Mixotoxodon and Piauhytherium became extinct at the end of the Late Pleistocene around 12,000 years ago as part of the Quaternary extinction event, along with most large mammals in the Americas. The extinction coincides with the arrival of the first humans to the Americas and they are suggested to have been a causal factor in the extinction.[2]
Orders and families
- Order Notoungulata - notoungulates
- Suborder Notioprogonia
- Family Henricosborniidae
- Family Notostylopidae
- Suborder Toxodontia
- Family Isotemnidae
- Family Leontiniidae
- Family Notohippidae
- Family Toxodontidae
- Family Homalodotheriidae
- Suborder Typotheria
- Family Archaeohyracidae
- Family Archaeopithecidae
- Family Campanorcidae
- Family Hegetotheriidae
- Family Interatheriidae
- Family Mesotheriidae
- Family Oldfieldthomasiidae
- Suborder Notioprogonia
References
- ↑ 1.0 1.1 Zimicz, Ana Natalia; Fernández, Mercedes; Bond, Mariano; Chornogubsky, Laura; Arnal, Michelle; Cárdenas, Magalí; Fernicola, Juan Carlos (November 2020). "Archaeogaia macachaae gen. et sp. nov., one of the oldest Notoungulata Roth, 1903 from the early-middle Paleocene Mealla Formation (Central Andes, Argentina) with insights into the Paleocene-Eocene south American biochronology" (in en). Journal of South American Earth Sciences 103: 102772. doi:10.1016/j.jsames.2020.102772. Bibcode: 2020JSAES.10302772Z.
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 2.6 2.7 Croft, Darin A.; Gelfo, Javier N.; López, Guillermo M. (2020-05-30). "Splendid Innovation: The Extinct South American Native Ungulates" (in en). Annual Review of Earth and Planetary Sciences 48 (1): 259–290. doi:10.1146/annurev-earth-072619-060126. ISSN 0084-6597. Bibcode: 2020AREPS..48..259C. https://www.annualreviews.org/doi/10.1146/annurev-earth-072619-060126.
- ↑ 3.0 3.1 Welker et al. 2015
- ↑ Buckley 2015
- ↑ 5.0 5.1 Westbury et al. 2017
- ↑ Cifelli 1993
- ↑ Missiaen et al. 2006
- ↑ Billet, Guillaume (December 2011). "Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters" (in en). Journal of Systematic Palaeontology 9 (4): 481–497. doi:10.1080/14772019.2010.528456. ISSN 1477-2019. http://www.tandfonline.com/doi/abs/10.1080/14772019.2010.528456.
- ↑ 9.0 9.1 Gomes Rodrigues, Helder; Herrel, Anthony; Billet, Guillaume (2017-01-31). "Ontogenetic and life history trait changes associated with convergent ecological specializations in extinct ungulate mammals" (in en). Proceedings of the National Academy of Sciences 114 (5): 1069–1074. doi:10.1073/pnas.1614029114. ISSN 0027-8424. PMID 28096389.
- ↑ 10.0 10.1 10.2 Cassini, Guillermo H.; Cerdeño, Esperanza; Villafañe, Amalia L.; Muñoz, Nahuel A. (2012-10-11), Vizcaíno, Sergio F.; Kay, Richard F.; Bargo, M. Susana, eds., "Paleobiology of Santacrucian native ungulates (Meridiungulata: Astrapotheria, Litopterna and Notoungulata)", Early Miocene Paleobiology in Patagonia (Cambridge University Press): pp. 243–286, doi:10.1017/cbo9780511667381.015, ISBN 978-0-511-66738-1, https://www.cambridge.org/core/product/identifier/9780511667381%23c19461-14-1/type/book_part, retrieved 2023-06-29
- ↑ Seckel, Lauren; Janis, Christine (December 2008). "Convergences in Scapula Morphology among Small Cursorial Mammals: An Osteological Correlate for Locomotory Specialization" (in en). Journal of Mammalian Evolution 15 (4): 261–279. doi:10.1007/s10914-008-9085-7. ISSN 1064-7554. http://link.springer.com/10.1007/s10914-008-9085-7.
- ↑ Croft, Darin A.; Lorente, Malena (2021-08-17). Smith, Thierry. ed. "No evidence for parallel evolution of cursorial limb adaptations among Neogene South American native ungulates (SANUs)" (in en). PLOS ONE 16 (8): e0256371. doi:10.1371/journal.pone.0256371. ISSN 1932-6203. PMID 34403434.
- ↑ 13.0 13.1 Seoane, Federico D.; Roig Juñent, Sergio; Cerdeño, Esperanza (2017-01-02). "Phylogeny and paleobiogeography of Hegetotheriidae (Mammalia, Notoungulata)" (in en). Journal of Vertebrate Paleontology 37 (1): e1278547. doi:10.1080/02724634.2017.1278547. ISSN 0272-4634. https://www.tandfonline.com/doi/full/10.1080/02724634.2017.1278547.
- ↑ Lundelius, Ernest L.; Bryant, Vaughn M.; Mandel, Rolfe; Thies, Kenneth J.; Thoms, Alston (January 2013). "The first occurrence of a toxodont (Mammalia, Notoungulata) in the United States" (in en). Journal of Vertebrate Paleontology 33 (1): 229–232. doi:10.1080/02724634.2012.711405. ISSN 0272-4634. http://www.tandfonline.com/doi/abs/10.1080/02724634.2012.711405.
- ↑ Fernández-Monescillo, Marcos; Martínez, Gastón; García López, Daniel; Frechen, Manfred; Romero-Lebrón, Eugenia; Krapovickas, Jerónimo M.; Haro, J. Augusto; Rodríguez, Pablo E. et al. (February 2023). "The last record of the last typotherid (Notoungulata, Mesotheriidae, Mesotherium cristatum) for the middle Pleistocene of the western Pampean region, Córdoba Province, Argentina, and its biostratigraphic implications" (in en). Quaternary Science Reviews 301: 107925. doi:10.1016/j.quascirev.2022.107925. https://linkinghub.elsevier.com/retrieve/pii/S027737912200556X.
Bibliography
- Billet, Guillaume (December 2011). "Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters". Journal of Systematic Palaeontology 9 (4): 481–97. doi:10.1080/14772019.2010.528456. OCLC 740994816.
- Buckley, M. (2015-04-01). "Ancient collagen reveals evolutionary history of the endemic South American 'ungulates'". Proceedings of the Royal Society B: Biological Sciences 282 (1806): 20142671. doi:10.1098/rspb.2014.2671. PMID 25833851.
- Cifelli, Richard L (1993). "The phylogeny of the native South American ungulates". in Szalay, F.S.; Novacek, M.J.; McKenna, M.C.. Mammal phylogeny: Placentals. 2. New York: Springer-Verlag. pp. 195–216. ISBN 0-387-97853-4. OCLC 715426850.
- Missiaen, P; Smith, T; Guo, DY; Bloch, JI; Gingerich, PD (August 2006). "Asian gliriform origin for arctostylopid mammals". Naturwissenschaften 93 (8): 407–11. doi:10.1007/s00114-006-0122-1. PMID 16865388. Bibcode: 2006NW.....93..407M. https://biblio.ugent.be/publication/353125.
- Roth, Santiago (1903). "Los Ungulados Sudamericanos". Anales del Museo de la Plata (Sección Paleontológica) 5: 1–36. OCLC 14012855.
- Welker, F.; Collins, M. J.; Thomas, J. A.; Wadsley, M.; Brace, S.; Cappellini, E.; Turvey, S. T.; Reguero, M. et al. (2015-03-18). "Ancient proteins resolve the evolutionary history of Darwin's South American ungulates". Nature 522 (7554): 81–84. doi:10.1038/nature14249. ISSN 0028-0836. PMID 25799987. Bibcode: 2015Natur.522...81W. http://eprints.whiterose.ac.uk/91438/1/Welker_postprint.docx.
- Westbury, M.; Baleka, S.; Barlow, A.; Hartmann, S.; Paijmans, J. L. A.; Kramarz, A.; Forasiepi, A. M.; Bond, M. et al. (2017-06-27). "A mitogenomic timetree for Darwin's enigmatic South American mammal Macrauchenia patachonica". Nature Communications 8: 15951. doi:10.1038/ncomms15951. PMID 28654082. Bibcode: 2017NatCo...815951W.
Further reading
- Carroll, Robert Lynn (1988). Vertebrate Paleontology and Evolution. New York: W.H. Freeman and Company. ISBN 9780716718222. OCLC 14967288.
- McKenna, M.C. (1975). "Toward a phylogenetic classification of the Mammalia". in Luckett, W.P.; Szalay, F.S.. Phylogeny of the primates: a multidisciplinary approach (Proceedings of WennerGren Symposium no. 61, Burg Wartenstein, Austria, July 6–14, 1974). New York: Plenum. pp. 21–46. doi:10.1007/978-1-4684-2166-8_2. ISBN 978-1-4684-2168-2. OCLC 1693999.
- McKenna, Malcolm C.; Bell, Susan K. (1997). Classification of Mammals Above the Species Level. New York: Columbia University Press. ISBN 0231110138. OCLC 37345734.
Wikidata ☰ Q131322 entry
 |  |

