Biology:Rsa RNA
Rsa RNAs are non-coding RNAs found in the bacterium Staphylococcus aureus. The shared name comes from their discovery, and does not imply homology. Bioinformatics scans identified the 16 Rsa RNA families named RsaA-K and RsaOA-OG.[1][2] Others, RsaOH-OX, were found thanks to an RNomic approach.[3] Although the RNAs showed varying expression patterns, many of the newly discovered RNAs were shown to be Hfq-independent and most carried a C-rich motif (UCCC).[1]
RsaA
Represses the translation of the transcriptional regulator MgrA by binding to its mRNA, enhances biofilm formation and decreases bacterial virulence.[4] Other mRNAs: including SsaA-like enzymes involved in peptidoglycan metabolism and the secreted anti-inflammatory FLIPr protein were validated as direct targets of RsaA.[5]
RsaE
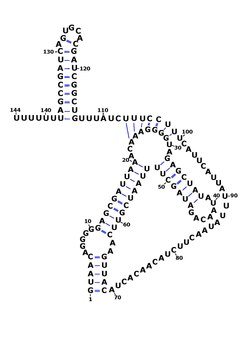
RsaE is found in other members of the genus Staphylococcus such as Staphylococcus epidermidis and Staphylococcus saprophyticus and is the only Rsa RNA to be found outside of this genus, in Macrococcus caseolyticus and Bacillus. In Bacillus subtilis, RsaE had previously been identified as ncr22.[8][9] RsaE is also consistently found downstream of PepF which codes for oligoendopeptidase F. The function of RsaE was discovered using gene knockout analysis and gene overexpression - it was found to regulate the expression of several enzymes involved in metabolism via antisense binding of their mRNA.[1][3]
RsaE was shown to be regulated by the presence of nitric oxide (NO). In Bacillus subtilis it controls expression of genes with functions related to oxidative stress and oxidation-reduction reactions and it was renamed RoxS (for related to oxidative stress).[10]
RsaF
In S.aureus species RsaF is located in the same intergenic region as RsaE and overlaps with 3′ end of RsaE by approximately 20bp. In contrast to RsaE, RsaF and its upstream gene have only been identified in S.aureus species.[1]
RsaK
RsaK is found in the leader sequence of glcA mRNA which encodes an enzyme involved in the glucose-specific phosphotransferase system. RsaK also contains a conserved ribonucleic antiterminator system, as recognised by GclT protein.[11]
RsaI
RsaOG[2] also renamed RsaI[1] is thought to fine-tune the regulation of toxin or invasion mechanisms in S. aureus via trans-acting mechanisms. Its secondary structure contains a pseudoknot formed between two highly conserved unpaired sequences.[6]
Expression patterns
RsaD, E H and I were found to be highly expressed in S. aureus. Expression levels of other Rsa RNAs varied under various environmental conditions, for example RsaC was induced by cold shock and RsaA is induced in response to osmotic stress.[1][2][3]
RsaE and RsaF genes overlap in S.aureus species but appear to have opposite expression patterns.[1] Transcriptional interference due to an overlap between a σA recognition motif and a potential σB binding site is proposed as a mechanism causing the differential expression of the two transcripts[1][12]
See also
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 "A search for small noncoding RNAs in Staphylococcus aureus reveals a conserved sequence motif for regulation". Nucleic Acids Research 37 (21): 7239–7257. November 2009. doi:10.1093/nar/gkp668. PMID 19786493.
- ↑ 2.0 2.1 2.2 "Single-pass classification of all noncoding sequences in a bacterial genome using phylogenetic profiles". Genome Research 19 (6): 1084–1092. June 2009. doi:10.1101/gr.089714.108. PMID 19237465.
- ↑ 3.0 3.1 3.2 "Experimental discovery of small RNAs in Staphylococcus aureus reveals a riboregulator of central metabolism". Nucleic Acids Research 38 (19): 6620–6636. October 2010. doi:10.1093/nar/gkq462. PMID 20511587.
- ↑ "A non-coding RNA promotes bacterial persistence and decreases virulence by regulating a regulator in Staphylococcus aureus". PLOS Pathogens 10 (3): e1003979. March 2014. doi:10.1371/journal.ppat.1003979. PMID 24651379.
- ↑ "The RNA targetome of Staphylococcus aureus non-coding RNA RsaA: impact on cell surface properties and defense mechanisms". Nucleic Acids Research 45 (11): 6746–6760. June 2017. doi:10.1093/nar/gkx219. PMID 28379505.
- ↑ 6.0 6.1 "RsaOG, a new staphylococcal family of highly transcribed non-coding RNA". RNA Biology 7 (2): 116–119. March 2010. doi:10.4161/rna.7.2.10925. PMID 20200491. http://www.landesbioscience.com/journals/rna/abstract.php?id=10925.
- ↑ "VARNA: Interactive drawing and editing of the RNA secondary structure". Bioinformatics 25 (15): 1974–1975. August 2009. doi:10.1093/bioinformatics/btp250. PMID 19398448.
- ↑ "The transcriptionally active regions in the genome of Bacillus subtilis". Molecular Microbiology 73 (6): 1043–1057. September 2009. doi:10.1111/j.1365-2958.2009.06830.x. PMID 19682248.
- ↑ "Identification of regulatory RNAs in Bacillus subtilis". Nucleic Acids Research 38 (19): 6637–6651. October 2010. doi:10.1093/nar/gkq454. PMID 20525796.
- ↑ "A nitric oxide regulated small RNA controls expression of genes involved in redox homeostasis in Bacillus subtilis". PLOS Genetics 11 (2): e1004957. February 2015. doi:10.1371/journal.pgen.1004957. PMID 25643072.
- ↑ "Specific interaction of the RNA-binding domain of the bacillus subtilis transcriptional antiterminator GlcT with its RNA target, RAT". Journal of Molecular Biology 293 (4): 795–805. November 1999. doi:10.1006/jmbi.1999.3176. PMID 10543968.
- ↑ "Transcriptional interference—a crash course". Trends in Genetics 21 (6): 339–345. June 2005. doi:10.1016/j.tig.2005.04.009. PMID 15922833.
Further reading
- "Small RNA genes expressed from Staphylococcus aureus genomic and pathogenicity islands with specific expression among pathogenic strains". Proceedings of the National Academy of Sciences of the United States of America 102 (40): 14249–14254. October 2005. doi:10.1073/pnas.0503838102. PMID 16183745.
- "Identification of bacterial small non-coding RNAs: experimental approaches". Current Opinion in Microbiology 10 (3): 257–261. June 2007. doi:10.1016/j.mib.2007.05.003. PMID 17553733.
- "Activation of alpha-toxin translation in Staphylococcus aureus by the trans-encoded antisense RNA, RNAIII". The EMBO Journal 14 (18): 4569–4577. September 1995. doi:10.1002/j.1460-2075.1995.tb00136.x. PMID 7556100.
- "Characterizing the effect of the Staphylococcus aureus virulence factor regulator, SarA, on log-phase mRNA half-lives". Journal of Bacteriology 188 (7): 2593–2603. April 2006. doi:10.1128/JB.188.7.2593-2603.2006. PMID 16547047.
- "Characterization of the Staphylococcus aureus heat shock, cold shock, stringent, and SOS responses and their effects on log-phase mRNA turnover". Journal of Bacteriology 188 (19): 6739–6756. October 2006. doi:10.1128/JB.00609-06. PMID 16980476.
- "Staphylococcus aureus RNAIII and the endoribonuclease III coordinately regulate spa gene expression". The EMBO Journal 24 (4): 824–835. February 2005. doi:10.1038/sj.emboj.7600572. PMID 15678100.
- "Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule". The EMBO Journal 12 (10): 3967–3975. October 1993. doi:10.1002/j.1460-2075.1993.tb06074.x. PMID 7691599.
- "No detectable effect of RNA-binding protein Hfq absence in Staphylococcus aureus". BMC Microbiology 7: 10. February 2007. doi:10.1186/1471-2180-7-10. PMID 17291347.
Gallery of Rsa RNA secondary structure images | |
|---|---|
|










 |  |

