Biology:Ampullae of Lorenzini

Ampullae of Lorenzini (Template:Singular: ampulla) are electroreceptors, sense organs able to detect electric fields. They form a network of mucus-filled pores in the skin of cartilaginous fish (sharks, rays, and chimaeras) and of basal bony fishes such as reedfish,[1] sturgeon,[2] and lungfish.[1] They are associated with and evolved from the mechanosensory lateral line organs of early vertebrates. Most bony fishes and terrestrial vertebrates have lost their ampullae of Lorenzini.
History
Ampullae were initially described by Marcello Malpighi and later given an exact description by the Italian physician and ichthyologist Stefano Lorenzini in 1679, though their function was unknown.[3] Electrophysiological experiments in the 20th century suggested a sensibility to temperature, mechanical pressure, and possibly salinity. In 1960 the ampullae were identified as specialized receptor organs for sensing electric fields.[4][5] One of the first descriptions of calcium-activated potassium channels was based on studies of the ampulla of Lorenzini in the skate.[6]
Evolution
Ampullae of Lorenzini are physically associated with and evolved from the mechanosensory lateral line organs of early vertebrates. Passive electroreception using ampullae is an ancestral trait in the vertebrates, meaning that it was present in their last common ancestor.[7] Ampullae of Lorenzini are present in cartilaginous fishes (sharks, rays, and chimaeras), lungfishes, bichirs, coelacanths, sturgeons, paddlefishes, aquatic salamanders, and caecilians. Ampullae of Lorenzini appear to have been lost early in the evolution of bony fishes and tetrapods, though the evidence for absence in many groups is incomplete and unsatisfactory.[7]
| Vertebrates |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Anatomy
Each ampulla is a bundle of sensory cells containing multiple nerve fibres in a sensory bulb (the endampulle) in a collagen sheath, and a gel-filled canal (the ampullengang) which opens to the surface by a pore in the skin. The gel is a glycoprotein-based substance with the same resistivity as seawater, and electrical properties similar to a semiconductor.[8][3][9]
Pores are concentrated in the skin around the snout and mouth of sharks and rays, as well as the anterior nasal flap, barbel, circumnarial fold and lower labial furrow.[10] Canal size typically corresponds to the body size of the animal but the number of ampullae remains the same. The canals of the ampullae of Lorenzini can be pored or non-pored. Non-pored canals do not interact with external fluid movement but serve a function as a tactile receptor to prevent interferences with foreign particles.[10]

Pores of ampullae of Lorenzini in snout of tiger shark
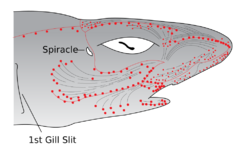
The electroreceptive ampullae of Lorenzini (red dots) evolved from the mechanosensory lateral line organs (gray lines) of early vertebrates.
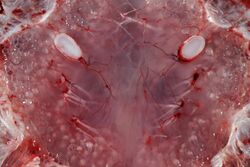
Inner view of ampullae of Lorenzini. The bulbs appear as many pale translucent spheres on either side of the midline.



Electroreception

The ampullae detect electric fields in the water, or more precisely the potential difference between the voltage at the skin pore and the voltage at the base of the electroreceptor cells.[11][12][6]
A positive pore stimulus decreases the rate of nerve activity coming from the electroreceptor cells, while a negative pore stimulus increases the rate. Each ampulla contains a single layer of receptor cells, separated by supporting cells. The cells are connected by apical tight junctions so that no current leaks between them. The apical faces of the receptor cells have a small surface area with a high concentration of voltage-dependent calcium channels (which trigger depolarisation) and calcium-activated potassium channels (for repolarisation afterwards).[13]
Because the canal wall has a very high resistance, all the voltage difference between the pore of the canal and the ampulla is dropped across the 50 micron-thick receptor epithelium. Because the basal membranes of the receptor cells have a lower resistance, most of the voltage is dropped across the excitable apical faces which are poised at the threshold. Inward calcium current across the receptor cells depolarises the basal faces, causing a large action potential, a wave of depolarisation followed by repolarisation (as in a nerve fibre). This triggers presynaptic calcium release and release of excitatory transmitter onto the afferent nerve fibres. These fibres signal the size of the detected electric field to the fish's brain.[14]
The ampulla contains large conductance calcium-activated potassium channels (BK channels). Sharks are much more sensitive to electric fields than electroreceptive freshwater fish, and indeed than any other animal, with a threshold of sensitivity as low as 5 nV/cm. The collagen jelly, a hydrogel, that fills the ampullae canals has one of the highest proton conductivity capabilities of any biological material. It contains keratan sulfate in 97% water, and has a conductivity of about 1.8 mS/cm.[14][11] All animals produce an electrical field caused by muscle contractions; electroreceptive fish may pick up weak electrical stimuli from the muscle contractions of their prey.[6]
The sawfish has more ampullary pores than any other cartilaginous fish, and is considered an electroreception specialist. Sawfish have ampullae of Lorenzini on their head, ventral and dorsal side of their rostrum leading to their gills, and on the dorsal side of their body.[15]
Magnetoreception
Ampullae of Lorenzini also contribute to the ability to receive geomagnetic information. As magnetic and electrical fields are related, magnetoreception via electromagnetic induction in the ampullae of Lorenzini is possible. Many cartilaginous fish respond to artificially generated magnetic fields in association with food rewards, demonstrating their capability. Magnetoreception may explain the ability of sharks and rays to form strict migratory patterns and to identify their geographic location.[16]
Temperature sense
The mucus-like substance inside the tubes was thought in 2003 perhaps to function as a thermoelectric semiconductor, transducing temperature changes into an electrical signal that the animal could use to detect temperature gradients.[17] A 2007 study appeared to disprove this.[18] The question remained open,[19] and in 2023 it was predicted that the ampullae of Lorenzini in sharks would be able to detect a temperature difference of 0.001 Kelvin (a thousandth of a degree). An artificial sensor using the same principle is able to detect a difference of 0.01 Kelvin.[20]
See also
- Knollenorgan – a non-homologous type of electroreceptor, found in mormyrid fishes
References
- ↑ 1.0 1.1 "Ultrastructure of the ampullary electroreceptors in lungfish and Brachiopterygii". Cell and Tissue Research 173 (1): 95–108. October 1976. doi:10.1007/BF00219268. PMID 991235.
- ↑ "Development of the lateral line system in the shovelnose sturgeon". Brain, Behavior and Evolution 64 (2): 70–84. 2004. doi:10.1159/000079117. PMID 15205543.
- ↑ 3.0 3.1 Baranes, Avi; Fishelson, Lev (6 January 1999). "Distribution, morphology, and cytology of ampullae of Lorenzini in the Oman shark, Iago omanensis (Triakidae), from the Gulf of Aqaba, Red Sea". The Anatomical Record 251 (251: 417–430): 417–430. doi:10.1002/(SICI)1097-0185(199808)251:4<417::AID-AR1>3.0.CO;2-P. PMID 9713980. https://onlinelibrary.wiley.com/doi/10.1002/(SICI)1097-0185(199808)251:4%3C417::AID-AR1%3E3.0.CO;2-P. Retrieved 21 March 2022.
- ↑ "Electrical sensitivity of the ampullae of Lorenzini". Nature 187 (4741): 957. September 1960. doi:10.1038/187957a0. PMID 13727039. Bibcode: 1960Natur.187..957M.
- ↑ "The response of the ampullae of Lorenzini of elasmobranchs to electrical stimulation". The Journal of Experimental Biology 39: 119–28. March 1962. doi:10.1242/jeb.39.1.119. PMID 14477490.
- ↑ 6.0 6.1 6.2 "The shark's electric sense.". Scientific American 297 (2): 74–81. August 2007. doi:10.1038/scientificamerican0807-74. PMID 17894175. Bibcode: 2007SciAm.297b..74F.
- ↑ 7.0 7.1 "The phylogenetic distribution of electroreception: Evidence for convergent evolution of a primitive vertebrate sense modality". Brain Research Reviews 6 (1): 25–46. 1983. doi:10.1016/0165-0173(83)90003-6. PMID 6616267. https://deepblue.lib.umich.edu/bitstream/2027.42/25137/1/0000573.pdf.
- ↑ "Semiconductor gel in shark sense organs?". Neuroscience Letters 426 (3): 166–70. October 2007. doi:10.1016/j.neulet.2007.08.064. PMID 17904741.
- ↑ "Ultrastructure of the ampullae of Lorenzini of Aptychotrema rostrata (Rhinobatidae)". Zoomorphology 128 (1): 45–52. 2009-02-01. doi:10.1007/s00435-008-0073-5. ISSN 1432-234X.
- ↑ 10.0 10.1 "Electroreceptive and mechanoreceptive anatomical specialisations in the epaulette shark (Hemiscyllium ocellatum)". PLOS One 7 (11): e49857. 2012. doi:10.1371/journal.pone.0049857. PMID 23226226. Bibcode: 2012PLoSO...749857W.
- ↑ 11.0 11.1 "Electroreception in early vertebrates: survey, evidence and new information.". Palaeontology 61 (3): 325–58. May 2018. doi:10.1111/pala.12346. https://phys.org/news/2018-02-sharks-animals-evolved-electroreception-theirprey.html.
- ↑ "Electroreception in marine fishes: chondrichthyans". Journal of Fish Biology 95 (1): 135–154. July 2019. doi:10.1111/jfb.14068. PMID 31169300.
- ↑ "Calcium-activated conductance in skate electroreceptors: current clamp experiments". The Journal of General Physiology 69 (2): 121–43. February 1977. doi:10.1085/jgp.69.2.121. PMID 190338.
- ↑ 14.0 14.1 "Molecular tuning of electroreception in sharks and skates". Nature 558 (7708): 122–126. June 2018. doi:10.1038/s41586-018-0160-9. PMID 29849147. Bibcode: 2018Natur.558..122B.
- ↑ "Sensory systems in sawfishes. 1. The ampullae of Lorenzini". Brain, Behavior and Evolution 78 (2): 139–49. 2011. doi:10.1159/000329515. PMID 21829004.
- ↑ "The yellow stingray (Urobatis jamaicensis) can discriminate the geomagnetic cues necessary for a bicoordinate magnetic map". Marine Biology 167 (10): 151. 2020-09-29. doi:10.1007/s00227-020-03763-1.
- ↑ "Sensing temperature without ion channels". Nature 421 (6922): 495–495. 30 January 2003. doi:10.1038/421495a. ISSN 0028-0836.
- ↑ "Semiconductor gel in shark sense organs?". Neuroscience Letters 426 (3): 166–170. 2007-10-22. doi:10.1016/j.neulet.2007.08.064. ISSN 0304-3940. PMID 17904741.
- ↑ "Temperature response in electrosensors and thermal voltages in electrolytes". Journal of Biological Physics 36 (2): 121–134. March 2010. doi:10.1007/s10867-009-9174-8. PMID 19760113.
- ↑ "Solid state ionics enabled ultra-sensitive detection of thermal trace with 0.001K resolution in deep sea". Nature Communications 14 (1). 12 January 2023. doi:10.1038/s41467-022-35682-8. ISSN 2041-1723.
| About fish |  | ||||
|---|---|---|---|---|---|
| Anatomy and physiology | |||||
| Sensory systems | |||||
| Reproduction | |||||
| Locomotion | |||||
| Other behaviour | |||||
| By habitat | |||||
| Other types | |||||
| Commercial |
| ||||
| Major groups | |||||
| Lists | |||||
 |  |

