Chemistry:Catechin
Catechin /ˈkætɪkɪn/ is a flavan-3-ol, a type of secondary metabolite providing antioxidant roles in plants. It belongs to the subgroup of polyphenols called flavonoids.
The name of the catechin chemical family derives from catechu, which is the tannic juice or boiled extract of Mimosa catechu (Acacia catechu L.f.).[1]
Chemistry

Catechin possesses two benzene rings (called the A and B rings) and a dihydropyran heterocycle (the C ring) with a hydroxyl group on carbon 3. The A ring is similar to a resorcinol moiety while the B ring is similar to a catechol moiety. There are two chiral centers on the molecule on carbons 2 and 3. Therefore, it has four diastereoisomers. Two of the isomers are in trans configuration and are called catechin and the other two are in cis configuration and are called epicatechin.
The most common catechin isomer is (+)-catechin. The other stereoisomer is (−)-catechin or ent-catechin. The most common epicatechin isomer is (−)-epicatechin (also known under the names L-epicatechin, epicatechol, (−)-epicatechol, L-acacatechin, L-epicatechol, epicatechin, 2,3-cis-epicatechin or (2R,3R)-(−)-epicatechin).
The different epimers can be separated using chiral column chromatography.[2]
Making reference to no particular isomer, the molecule can just be called catechin. Mixtures of the different enantiomers can be called (±)-catechin or DL-catechin and (±)-epicatechin or DL-epicatechin.
Catechin and epicatechin are the building blocks of the proanthocyanidins, a type of condensed tannin.
- Diastereoisomers gallery
-
 (+)-catechin (2R,3S)
(+)-catechin (2R,3S) -
 (−)-catechin (2S,3R)
(−)-catechin (2S,3R) -
 (−)-epicatechin (2R,3R)
(−)-epicatechin (2R,3R) -
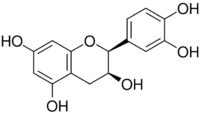 (+)-epicatechin (2S,3S)
(+)-epicatechin (2S,3S)
-Catechin.png)

-Epicatechin.svg)
-epicatechin.svg)

Moreover, the flexibility of the C-ring allows for two conformation isomers, putting the B-ring either in a pseudoequatorial position (E conformer) or in a pseudoaxial position (A conformer). Studies confirmed that (+)-catechin adopts a mixture of A- and E-conformers in aqueous solution and their conformational equilibrium has been evaluated to be 33:67.[3]
As flavonoids, catechins can act as antioxidants when in high concentration in vitro, but compared with other flavonoids, their antioxidant potential is low.[4] The ability to quench singlet oxygen seems to be in relation with the chemical structure of catechin, with the presence of the catechol moiety on ring B and the presence of a hydroxyl group activating the double bond on ring C.[5]
Oxidation
Electrochemical experiments show that (+)-catechin oxidation mechanism proceeds in sequential steps, related with the catechol and resorcinol groups and the oxidation is pH-dependent. The oxidation of the catechol 3′,4′-dihydroxyl electron-donating groups occurs first, at very low positive potentials, and is a reversible reaction. The hydroxyl groups of the resorcinol moiety oxidised afterwards were shown to undergo an irreversible oxidation reaction.[6]
The laccase–ABTS system oxidizes (+)-catechin to oligomeric products[7] of which proanthocyanidin A2 is a dimer.
Spectral data

| UV-Vis | |
|---|---|
| Lambda-max: | 276 nm |
| Extinction coefficient (log ε) | 4.01 |
| IR | |
| Major absorption bands | 1600 cm−1(benzene rings) |
| NMR | |
| Proton NMR
|
δ : 2.49 (1H, dd, J = 16.0, 8.6 Hz, H-4a), |
| Carbon-13 NMR | |
| Other NMR data | |
| MS | |
| Masses of main fragments |
ESI-MS [M+H]+ m/z : 291.0
|
Natural occurrences
(+)-Catechin and (−)-epicatechin as well as their gallic acid conjugates are ubiquitous constituents of vascular plants, and frequent components of traditional herbal remedies, such as Uncaria rhynchophylla. The two isomers are mostly found as cacao and tea constituents, as well as in Vitis vinifera grapes.[9][10][11]
In food
The main dietary sources of catechins in Europe and the United States are tea and pome fruits.[12][13]
Catechins and epicatechins are found in cocoa,[14] which, according to one database, has the highest content (108 mg/100 g) of catechins among foods analyzed, followed by prune juice (25 mg/100 ml) and broad bean pod (16 mg/100 g).[15] Açaí oil, obtained from the fruit of the açaí palm (Euterpe oleracea), contains (+)-catechins (67 mg/kg).[16]
Catechins are diverse among foods,[15] from peaches[17] to green tea and vinegar.[15][18] Catechins are found in barley grain, where they are the main phenolic compound responsible for dough discoloration.[19] The taste associated with monomeric (+)-catechin or (−)-epicatechin is described as slightly astringent, but not bitter.[20]
Metabolism
Biosynthesis
File:Biosynthesis of 4-hydroxycinnamoyl-CoA.png
The biosynthesis of catechin begins with ma 4-hydroxycinnamoyl CoA starter unit which undergoes chain extension by the addition of three malonyl-CoAs through a PKSIII pathway. 4-Hydroxycinnamoyl CoA is biosynthesized from L-phenylalanine through the Shikimate pathway. L-Phenylalanine is first deaminated by phenylalanine ammonia lyase (PAL) forming cinnamic acid which is then oxidized to 4-hydroxycinnamic acid by cinnamate 4-hydroxylase. Chalcone synthase then catalyzes the condensation of 4-hydroxycinnamoyl CoA and three molecules of malonyl-CoA to form chalcone. Chalcone is then isomerized to naringenin by chalcone isomerase which is oxidized to eriodictyol by flavonoid 3′-hydroxylase and further oxidized to taxifolin by flavanone 3-hydroxylase. Taxifolin is then reduced by dihydroflavanol 4-reductase and leucoanthocyanidin reductase to yield catechin. The biosynthesis of catechin is shown below[21][22][23]
Leucocyanidin reductase (LCR) uses 2,3-trans-3,4-cis-leucocyanidin to produce (+)-catechin and is the first enzyme in the proanthocyanidin (PA) specific pathway. Its activity has been measured in leaves, flowers, and seeds of the legumes Medicago sativa, Lotus japonicus, Lotus uliginosus, Hedysarum sulfurescens, and Robinia pseudoacacia.[24] The enzyme is also present in Vitis vinifera (grape).[25]
File:Biosynthesis of catechin.png
Biodegradation
Catechin oxygenase, a key enzyme in the degradation of catechin, is present in fungi and bacteria.[26]
Among bacteria, degradation of (+)-catechin can be achieved by Acinetobacter calcoaceticus. Catechin is metabolized to protocatechuic acid (PCA) and phloroglucinol carboxylic acid (PGCA).[27] It is also degraded by Bradyrhizobium japonicum. Phloroglucinol carboxylic acid is further decarboxylated to phloroglucinol, which is dehydroxylated to resorcinol. Resorcinol is hydroxylated to hydroxyquinol. Protocatechuic acid and hydroxyquinol undergo intradiol cleavage through protocatechuate 3,4-dioxygenase and hydroxyquinol 1,2-dioxygenase to form β-carboxy-cis,cis-muconic acid and maleyl acetate.[28]
Among fungi, degradation of catechin can be achieved by Chaetomium cupreum.[29]
Metabolism in humans

-epicatechin_metabolism_in_humans_as_a_function_of_time_post-oral_intake.jpg)
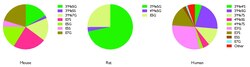
Catechins are metabolised upon uptake from the gastrointestinal tract, in particular the jejunum,[31] and in the liver, resulting in so-called structurally related epicatechin metabolites (SREM).[32] The main metabolic pathways for SREMs are glucuronidation, sulfation and methylation of the catechol group by catechol-O-methyl transferase, with only small amounts detected in plasma.[33][30] The majority of dietary catechins are however metabolised by the colonic microbiome to gamma-valerolactones and hippuric acids which undergo further biotransformation, glucuronidation, sulfation and methylation in the liver.[33]
The stereochemical configuration of catechins has a strong impact on their uptake and metabolism as uptake is highest for (−)-epicatechin and lowest for (−)-catechin.[34]
Biotransformation
Biotransformation of (+)-catechin into taxifolin by a two-step oxidation can be achieved by Burkholderia sp.[35]
(+)-Catechin and (−)-epicatechin are transformed by the endophytic filamentous fungus Diaporthe sp. into the 3,4-cis-dihydroxyflavan derivatives, (+)-(2R,3S,4S)-3,4,5,7,3′,4′-hexahydroxyflavan (leucocyanidin) and (−)-(2R,3R,4R)-3,4,5,7,3′,4′-hexahydroxyflavan, respectively, whereas (−)-catechin and (+)-epicatechin with a (2S)-phenyl group resisted the biooxidation.[36]
Leucoanthocyanidin reductase (LAR) uses (2R,3S)-catechin, NADP+ and H2O to produce 2,3-trans-3,4-cis-leucocyanidin, NADPH, and H+. Its gene expression has been studied in developing grape berries and grapevine leaves.[37]
Glycosides
- (2R,3S)-Catechin-7-O-β-D-glucopyranoside can be isolated from barley (Hordeum vulgare L.) and malt.[38]
- Epigeoside (catechin-3-O-α-L-rhamnopyranosyl-(1–4)-β-D-glucopyranosyl-(1–6)-β-D-glucopyranoside) can be isolated from the rhizomes of Epigynum auritum.[39]
Research
Vascular function
Only limited evidence from dietary studies indicates that catechins may affect endothelium-dependent vasodilation which could contribute to normal blood flow regulation in humans.[40][41] Green tea catechins may improve blood pressure, especially when systolic blood pressure is above 130 mmHg.[42][43]
Due to extensive metabolism during digestion, the fate and activity of catechin metabolites responsible for this effect on blood vessels, as well as the actual mode of action, are unknown.[33][44]
Adverse events
Catechin and its metabolites can bind tightly to red blood cells and thereby induce the development of autoantibodies, resulting in haemolytic anaemia and renal failure.[45] This resulted in the withdrawal of the catechin-containing drug Catergen, used to treat viral hepatitis,[46] from market in 1985.[47]
Catechins from green tea can be hepatotoxic[48] and the European Food Safety Authority has recommended not to exceed 800 mg per day.[49]
Other
One limited meta-analysis showed that increasing consumption of green tea and its catechins to seven cups per day provided a small reduction in prostate cancer.[50] Nanoparticle methods are under preliminary research as potential delivery systems of catechins.[51]
Botanical effects
Catechins released into the ground by some plants may hinder the growth of their neighbors, a form of allelopathy.[52] Centaurea maculosa, the spotted knapweed often studied for this behavior, releases catechin isomers into the ground through its roots, potentially having effects as an antibiotic or herbicide. One hypothesis is that it causes a reactive oxygen species wave through the target plant's root to kill root cells by apoptosis.[53] Most plants in the European ecosystem have defenses against catechin, but few plants are protected against it in the North American ecosystem where Centaurea maculosa is an invasive, uncontrolled weed.[52]
Catechin acts as an infection-inhibiting factor in strawberry leaves.[54] Epicatechin and catechin may prevent coffee berry disease by inhibiting appressorial melanization of Colletotrichum kahawae.[55]
References
- ↑ "Cutch and catechu plant origin". Food and Agriculture Organization of the United Nations. 5 November 2011. http://www.fao.org/docrep/V8879E/v8879e05.htm.
- ↑ "Determination of catechin diastereomers from the leaves of Byrsonima species using chiral HPLC-PAD-CD". Chirality 22 (8): 726–733. August 2010. doi:10.1002/chir.20824. PMID 20143413.
- ↑ "Investigation of the complexation of (+)-catechin by beta-cyclodextrin by a combination of NMR, microcalorimetry and molecular modeling techniques". Organic & Biomolecular Chemistry 1 (14): 2590–2595. July 2003. doi:10.1039/B302935M. PMID 12956082.
- ↑ "Flavonoids as antioxidants". Journal of Natural Products 63 (7): 1035–1042. July 2000. doi:10.1021/np9904509. PMID 10924197. Bibcode: 2000JNAtP..63.1035P.
- ↑ "Antioxidant activity of flavonoids: efficiency of singlet oxygen (1Δg) quenching". Journal of Photochemistry and Photobiology B: Biology 19 (3): 205–215. August 1993. doi:10.1016/1011-1344(93)87086-3. PMID 8229463.
- ↑ "Catechin electrochemical oxidation mechanisms". Analytica Chimica Acta 518 (1–2): 109–115. 2004. doi:10.1016/j.aca.2004.05.038. Bibcode: 2004AcAC..518..109J.
- ↑ "The laccase/ABTS system oxidizes (+)-catechin to oligomeric products". Enzyme and Microbial Technology 40 (5): 1272–1279. April 2007. doi:10.1016/j.enzmictec.2006.09.018.
- ↑ "Neural cell protective compounds isolated from Phoenix hanceana var. formosana". Phytochemistry 70 (9): 1173–1181. June 2009. doi:10.1016/j.phytochem.2009.06.006. PMID 19628235. Bibcode: 2009PChem..70.1173L.
- ↑ "Optimization of supercritical fluid consecutive extractions of fatty acids and polyphenols from Vitis vinifera grape wastes". Journal of Food Science 80 (1): E101–E107. January 2015. doi:10.1111/1750-3841.12715. PMID 25471637.
- ↑ "The Catechin of the Cacao Bean". Journal of the American Chemical Society 54 (5): 1913–1917. 1932. doi:10.1021/ja01344a026. Bibcode: 1932JAChS..54.1913F.
- ↑ "Michiyo Tsujimura (1888–1969)". http://archives.cf.ocha.ac.jp/en/researcher/tsujimura_michiyo.html.
- ↑ "Estimated dietary flavonoid intake and major food sources of U.S. adults". The Journal of Nutrition 137 (5): 1244–1252. May 2007. doi:10.1093/jn/137.5.1244. PMID 17449588.
- ↑ "Flavonoid intake in European adults (18 to 64 years)". PLOS ONE 10 (5). 2015. doi:10.1371/journal.pone.0128132. PMID 26010916. Bibcode: 2015PLoSO..1028132V.
- ↑ "Cocoa flavanols – measurement, bioavailability and bioactivity". Asia Pacific Journal of Clinical Nutrition 17 (Suppl. 1): 280–283. 2008. PMID 18296356. http://apjcn.nhri.org.tw/server/APJCN/17%20Suppl%201//280.pdf.
- ↑ 15.0 15.1 15.2 "Polyphenols in green tea infusion". Phenol-Explorer, v3.5. 2014. http://phenol-explorer.eu/contents/food/29.
- ↑ "Chemical composition, antioxidant properties, and thermal stability of a phytochemical enriched oil from Açaí (Euterpe oleracea Mart.)". Journal of Agricultural and Food Chemistry 56 (12): 4631–4636. June 2008. doi:10.1021/jf800161u. PMID 18522407. Bibcode: 2008JAFC...56.4631P.
- ↑ "Browning Potential, Phenolic Composition, and Polyphenoloxidase Activity of Buffer Extracts of Peach and Nectarine Skin Tissue". Journal of the American Society for Horticultural Science 120 (5): 835–838. 1995. doi:10.21273/JASHS.120.5.835.
- ↑ "Analysis of polyphenolic compounds of different vinegar samples". Zeitschrift für Lebensmittel-Untersuchung und -Forschung 199 (1): 29–31. 1994. doi:10.1007/BF01192948.
- ↑ "Phenolic compounds of barley grain and their implication in food product discoloration". Journal of Agricultural and Food Chemistry 54 (26): 9978–9984. December 2006. doi:10.1021/jf060974w. PMID 17177530. Bibcode: 2006JAFC...54.9978Q.
- ↑ Kielhorn, S.; Thorngate, J. H. III (1999). "Oral sensations associated with the flavan-3-ols (+)-catechin and (−)-epicatechin". Food Quality and Preference 10 (2): 109–116. doi:10.1016/S0950-3293(98)00049-4.
- ↑ "Molecular regulation of catechins biosynthesis in tea [Camellia sinensis (L.) O. Kuntze]". Gene 495 (2): 205–210. March 2012. doi:10.1016/j.gene.2011.12.029. PMID 22226811.
- ↑ "Flavonoid biosynthesis in the tea plant Camellia sinensis: properties of enzymes of the prominent epicatechin and catechin pathways". Archives of Biochemistry and Biophysics 431 (1): 22–30. November 2004. doi:10.1016/j.abb.2004.08.003. PMID 15464723.
- ↑ Medicinal Natural Products: A Biosynthetic Approach (3rd ed.). UK: John Wiley & Sons. 2009. ISBN 978-0-470-74167-2.
- ↑ "Leucocyanidin Reductase Activity and Accumulation of Proanthocyanidins in Developing Legume Tissues". American Journal of Botany 84 (4): 494–503. April 1997. doi:10.2307/2446026.
- ↑ "Crystal structure and catalytic mechanism of leucoanthocyanidin reductase from Vitis vinifera". Journal of Molecular Biology 397 (4): 1079–1091. April 2010. doi:10.1016/j.jmb.2010.02.002. PMID 20138891.
- ↑ Arunachalam, M.; Mohan Raj, M.; Mohan, N.; Mahadevan, A. (2003). "Biodegradation of Catechin". Proceedings of the Indian National Science Academy B69 (4): 353–370. http://www.new.dli.ernet.in/rawdataupload/upload/insa/INSA_1/20008a2f_353.pdf.
- ↑ "Degradation of (+)-catechin by Acinetobacter calcoaceticus MTC 127". Biochimica et Biophysica Acta (BBA) - General Subjects 1621 (3): 261–265. June 2003. doi:10.1016/S0304-4165(03)00077-1. PMID 12787923.
- ↑ "Degradation of catechin by Bradyrhizobium japonicum". Biodegradation 8 (3): 159–165. 1997. doi:10.1023/A:1008254812074.
- ↑ "Degradation of catechin and purification and partial characterization of catechin oxygenase from Chaetomium cupreum". World Journal of Microbiology & Biotechnology 9 (1): 37–44. January 1993. doi:10.1007/BF00656513. PMID 24419836.
- ↑ 30.0 30.1 30.2 30.3 "The metabolome of [2-14C(−)-epicatechin in humans: implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives"]. Scientific Reports 6. July 2016. doi:10.1038/srep29034. PMID 27363516. Bibcode: 2016NatSR...629034O.
- ↑ "Intestinal absorption, metabolism, and excretion of (−)-epicatechin in healthy humans assessed by using an intestinal perfusion technique". The American Journal of Clinical Nutrition 98 (4): 924–933. October 2013. doi:10.3945/ajcn.113.065789. PMID 23864538.
- ↑ "Structurally related (−)-epicatechin metabolites in humans: assessment using de novo chemically synthesized authentic standards". Free Radical Biology & Medicine 52 (8): 1403–1412. April 2012. doi:10.1016/j.freeradbiomed.2011.12.010. PMID 22240152.
- ↑ 33.0 33.1 33.2 "Flavonoids". Linus Pauling Institute, Oregon State University, Corvallis. 2016. http://lpi.oregonstate.edu/mic/dietary-factors/phytochemicals/flavonoids.
- ↑ "The stereochemical configuration of flavanols influences the level and metabolism of flavanols in humans and their biological activity in vivo". Free Radical Biology & Medicine 50 (2): 237–244. January 2011. doi:10.1016/j.freeradbiomed.2010.11.005. PMID 21074608.
- ↑ "Biotransformation of (+)-catechin into taxifolin by a two-step oxidation: primary stage of (+)-catechin metabolism by a novel (+)-catechin-degrading bacteria, Burkholderia sp. KTC-1, isolated from tropical peat". Biochemical and Biophysical Research Communications 366 (2): 414–419. February 2008. doi:10.1016/j.bbrc.2007.11.157. PMID 18068670. Bibcode: 2008BBRC..366..414M.
- ↑ "Biooxidation of (+)-catechin and (−)-epicatechin into 3,4-dihydroxyflavan derivatives by the endophytic fungus Diaporthe sp. isolated from a tea plant". Chemical & Pharmaceutical Bulletin 53 (7): 866–867. July 2005. doi:10.1248/cpb.53.866. PMID 15997157.
- ↑ "Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves". Plant Physiology 139 (2): 652–663. October 2005. doi:10.1104/pp.105.064238. PMID 16169968.
- ↑ "Identification of a new flavanol glucoside from barley (Hordeum vulgare L.) and malt". European Food Research and Technology 214 (5): 388–393. 2002. doi:10.1007/s00217-002-0498-x.
- ↑ "[Study on glycosidal constituents from Epigynum auritum]" (in zh). Yao Xue Xue Bao (Acta Pharmaceutica Sinica) 26 (11): 841–845. 1991. PMID 1823978.
- ↑ "Effects of chocolate, cocoa, and flavan-3-ols on cardiovascular health: a systematic review and meta-analysis of randomized trials". The American Journal of Clinical Nutrition 95 (3): 740–751. March 2012. doi:10.3945/ajcn.111.023457. PMID 22301923.
- ↑ "Epicatechin ingested via cocoa products reduces blood pressure in humans: a nonlinear regression model with a Bayesian approach". The American Journal of Clinical Nutrition 95 (6): 1365–1377. June 2012. doi:10.3945/ajcn.111.029330. PMID 22552030.
- ↑ "Green tea catechins and blood pressure: a systematic review and meta-analysis of randomised controlled trials". European Journal of Nutrition 53 (6): 1299–1311. September 2014. doi:10.1007/s00394-014-0720-1. PMID 24861099.
- ↑ "The Cardiovascular Effects of Cocoa Polyphenols—An Overview". Diseases 4 (4): 39. December 2016. doi:10.3390/diseases4040039. PMID 28933419.
- ↑ "(−)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans". Proceedings of the National Academy of Sciences of the United States of America 103 (4): 1024–1029. January 2006. doi:10.1073/pnas.0510168103. PMID 16418281. Bibcode: 2006PNAS..103.1024S.
- ↑ "Toxicology and Safety of Flavonoids". Methods of Analysis, Preclinical and Clinical Pharmacokinetics, Safety, and Toxicology. John Wiley & Son. 2013. p. 257. ISBN 978-0-470-57871-1.
- ↑ Okolicsányi, Lajos; Csomós, Géza; Crepaldi, Gaetano, eds (1987). Assessment and Management of Hepatobiliary Disease. Berlin: Springer-Verlag. p. 371. doi:10.1007/978-3-642-72631-6. ISBN 978-3-642-72631-6.
- ↑ "Ruhen der Zulassung für Catergen" (in de). Deutsches Ärzteblatt 82 (38): 2706. https://www.aerzteblatt.de/pdf/82/38/a2706.pdf.
- ↑ Health Canada (2017-11-15). "Summary Safety Review – Green tea extract-containing natural health products – Assessing the potential risk of liver injury (hepatotoxicity)". https://www.canada.ca/en/health-canada/services/drugs-health-products/medeffect-canada/safety-reviews/green-tea-extract-containing-natural-health-products-assessing-potential-risk-liver-injury.html.
- ↑ "Scientific opinion on the safety of green tea catechins". EFSA Journal 16 (4): e05239. April 2018. doi:10.2903/j.efsa.2018.5239. PMID 32625874.
- ↑ "Green tea and the risk of prostate cancer: A systematic review and meta-analysis". Medicine 96 (13). March 2017. doi:10.1097/MD.0000000000006426. PMID 28353571.
- ↑ "Nano- and micro-particles for delivery of catechins: Physical and biological performance". Critical Reviews in Food Science and Nutrition 59 (10): 1563–1579. 2018. doi:10.1080/10408398.2017.1422110. PMID 29345975.
- ↑ 52.0 52.1 "Secondary Metabolites and Allelopathy in Plant Invasions: A Case Study of Centaurea maculosa". Plant Physiology and Development (6th ed.). Sinauer Associates. 2006. http://6e.plantphys.net/essay23.07.html.
- ↑ "Allelopathy and exotic plant invasion: from molecules and genes to species interactions". Science 301 (5638): 1377–1380. September 2003. doi:10.1126/science.1083245. PMID 12958360. Bibcode: 2003Sci...301.1377B.
- ↑ "(+)-Catechin acts as an infection-inhibiting factor in strawberry leaf". Phytopathology 90 (6): 595–600. June 2000. doi:10.1094/PHYTO.2000.90.6.595. PMID 18944538. Bibcode: 2000PhPat..90..595Y.
- ↑ "Epicatechin and catechin may prevent coffee berry disease by inhibition of appressorial melanization of Colletotrichum kahawae". Biotechnology Letters 28 (20): 1637–1640. October 2006. doi:10.1007/s10529-006-9135-2. PMID 16955359.
External links
 |  |
