Biology:N-acetylglucosamine-6-phosphate deacetylase
| N-acetylglucosamine-6-phosphate deacetylase in Mycobacterium smegmatis | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| EC number | 3.5.1.25 | ||||||||
| CAS number | 9027-50-3 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
In enzymology, N-acetylglucosamine-6-phosphate deacetylase (EC 3.5.1.25), also known as GlcNAc-6-phosphate deacetylase or NagA, is an enzyme that catalyzes the deacetylation of N-acetylglucosamine-6-phosphate (GlcNAc-6-P) to glucosamine-6-phosphate (GlcN-6-P):
- H2O + N-acetyl-D-glucosamine 6-phosphate acetate + D-glucosamine 6-phosphate[1]
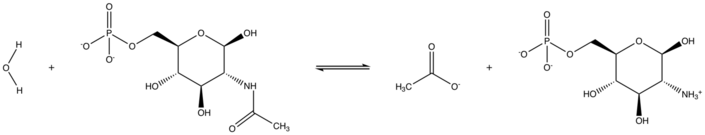
NagA Reaction

GlcNAc-6-phosphate deacetylase is encoded by the gene NagA.[2]
This enzyme belongs to the amidohydrolase superfamily.[3] Amidohydrolases are a type of hydrolase that acts upon amide bonds. All members of the amidohydrolase family employ a TIM barrel structure, and a vast majority of members are metalloenzymes.[4] The family of enzymes is important in amino acid and nucleotide metabolism as well as biodegradation of agricultural and industrial compounds. NagA participates in amino-sugar metabolism, specifically in the biosynthesis of amino-sugar-nucleotides.[5]
Structure
NagA is a homodimeric enzyme with two domains in each dimer of the structure.[6] Each domain I comprises a (β/α)8 - barrel structural fold, also known as a TIM barrel, and contains an active site of the enzyme. Each active site consists of the catalytic site of the enzyme and the metal-binding site that are involved in substrate and metal co-factor recognition, respectively. Domain I also forms the dimeric interface with domain I of the neighboring subunit.[6] The smaller second domain of NagA enzymes comprises a β-barrel, which potentially acts to stabilize the enzyme.[6] While all members of the amidohydrolase superfamily employ a TIM-barrel structural fold, NagA in Escherichia coli (EcNagA) has a pseudo-TIM barrel enclosing the funnel-like catalytic site of the enzyme.[7] The dimer structure of NagA is considered crucial for the activity and thermostability of the enzyme.[8]

Metal-binding site
Amidohydrolase enzymes can bind one, two, or three metal atoms in the active site. These metals can include Zn2+, Co2+, Fe2+, Cd2+, and others.[1] EcNagA contains a mononuclear metal-binding site with a Zn2+ ion;[3] in addition, EcNagA shows a phosphate ion bound at the metal-binding site.[7] Unlike EcNagA, NagA of Mycobacterium smegmatis (MSNagA) and Bacillus subtilis (BsNagA) have binuclear metal-binding sites. MSNagA has two divalent metal ions located in each active site, which are both required for efficient catalysis and structural stability.[6] While most other bacteria species use Zn as their metal co-factor, BsNagA utilizes iron as the predominant metal in the metal-binding site.[9]
Catalytic-binding site
Most of the active site residues of EcNagA and BsNagA are conserved and share similar structural positions. A notable difference between mycobacterial NagA enzymes and NagA enzymes from other bacterial species is the presence of a cysteine at position 131. Other bacterial species have a lysine residue at this position. This cysteine is located in the flexible loop, which prevents the physiological substrate from binding.[6]
Mechanism
The catalytic mechanism for NagA enzymes proposed utilizes nucleophilic attack via a metal-coordinated water molecule or hydroxide ion. The mechanism proceeds via a strictly conserved active-site aspartic acid residue (Asp-273) that acts initially as a base to activate the hydrolytic water molecule in order to attack the carbonyl group of the substrate.[3] Asp-273 then acts as an acid to protonate the amine leaving group. One proposed mechanism using the BsNagA and its two iron co-factors in the metal-binding site demonstrates the nucleophilic attack by an Fe-bridged hydroxide and then the stabilization of the carbonyl oxygen by one of the two Fe atoms.[9]
Biological Function

NagA is located in the cytoplasm of the cell. N-acetylglucosamine (GlcNAc) enters the cell as part of the breakdown of the cell wall. GlcNAc, a monosaccharide and derivative of glucose, is part of a biopolymer in the bacterial cell wall. This biopolymer forms a layered structure called peptidoglycan (PG). GlcNAc is then converted into GlcNAc-6-P by the enzyme NagE.[10] This substrate is then deacetylated into acetate and GlcN-6-P by NagA.[11] NagA is important for the production of GlcN-6-P, which is then used in two main pathways: PG recycling pathway and the glycolysis pathway.
PG recycling pathway
In the PG Recycling pathway, once GlcNAc-6-P is metabolized by NagA, its product, GlcN-6-P, can then be converted to GlcN-1-P by the enzyme GlmM, followed by reacetylation and reaction with UTP by GlmU to form UDP-GlcNAc.[10][11] UDP-GlcNAc is the end product of this pathway, which is then used to make glycosaminoglycans, proteoglycans, and glycolipids, which are all necessary in order to replenish PG for the cell wall.[12] PG recycling is necessary for bacterial cells in order to ensure bacteria growth and prevent cell lysis.[13]
Glycolysis pathway
Instead of entering the PG recycling pathway, GlcN-6-P can be converted into fructose-6-phosphate by NagB. This reaction is reversible by the enzyme GlmS,[10][11] an amidotransferase.[13] The produced fructose-6-phosphate then enters the glycolysis pathway. Glycolysis catalyzes the production of pyruvate, leading to the citric acid cycle and allowing for the production of amino acids.[14] GlcN-6-P and fructose-6-phosphate act as allosteric regulators of NagA, inhibiting further deacetylation of GlcNAc-6-P.[15]
Disease relevance
NagA is a potential drug target of Mycobacterium tuberculosis (Mtb). Eliminating NagA produces high levels of the allosteric activator GlcNAc-6-P,[2] which prevents the production of GlcN-6-P in order to proceed with the PG recycling pathway. NagA is, therefore, at a crucial metabolic chokepoint in Mtb,[16] representing the key enzymatic step in the generation of essential amino-sugar precursors. These precursors are required for Mtb cell wall biosynthesis and influence the PG recycling pathway. Additionally, the presence of cysteine in MSNagA's active site may represent a unique exploitative target in Mtb therapeutics.[6]
Structural studies
As of early 2019, 11 structures have been solved for this class of enzymes, with PDB accession codes 1O12, 1UN7, 1YMY, 1YRR, 2P50, 2P53, 6FV3, 6FV4, 3EGJ, 3IV8, and 2VHL.
Nomenclature
The systematic name of this enzyme class is N-acetyl-D-glucosamine-6-phosphate amidohydrolase. Other names in common use include acetylglucosamine phosphate deacetylase, acetylaminodeoxyglucosephosphate acetylhydrolase, and 2-acetamido-2-deoxy-D-glucose-6-phosphate amidohydrolase.[15]
References
- ↑ 1.0 1.1 "nagA - N-acetylglucosamine-6-phosphate deacetylase - Escherichia coli (strain K12) - nagA gene & protein". https://www.uniprot.org/uniprot/P0AF18.
- ↑ 2.0 2.1 "Allosteric regulation of glucosamine-6-phosphate deaminase (NagB) and growth of Escherichia coli on glucosamine". Journal of Bacteriology 191 (20): 6401–7. October 2009. doi:10.1128/JB.00633-09. PMID 19700525.
- ↑ 3.0 3.1 3.2 "N-Acetyl-D-glucosamine-6-phosphate deacetylase: substrate activation via a single divalent metal ion". Biochemistry 46 (27): 7942–52. July 2007. doi:10.1021/bi700543x. PMID 17567047.
- ↑ Liu, Aimin; Huo, Lu (2014-08-15), John Wiley & Sons Ltd, ed., "Amidohydrolase Superfamily", eLS (John Wiley & Sons, Ltd), doi:10.1002/9780470015902.a0020546.pub2, ISBN 9780470015902
- ↑ "N-acetylglucosamine 6-phosphate deacetylase (nagA) is required for N-acetyl glucosamine assimilation in Gluconacetobacter xylinus". PLOS ONE 6 (6): e18099. 2011-06-02. doi:10.1371/journal.pone.0018099. PMID 21655093. Bibcode: 2011PLoSO...618099Y.
- ↑ 6.0 6.1 6.2 6.3 6.4 6.5 "Mycobacterium tuberculosis N-acetylglucosamine-6-phosphate deacetylase (NagA)". The Journal of Biological Chemistry 293 (25): 9770–9783. June 2018. doi:10.1074/jbc.RA118.002597. PMID 29728457.
- ↑ 7.0 7.1 "Structural analysis of N-acetylglucosamine-6-phosphate deacetylase apoenzyme from Escherichia coli". Journal of Molecular Biology 359 (2): 308–21. June 2006. doi:10.1016/j.jmb.2006.03.024. PMID 16630633.
- ↑ "The structure of hyperthermophilic β-N-acetylglucosaminidase reveals a novel dimer architecture associated with the active site". The FEBS Journal 281 (22): 5092–103. November 2014. doi:10.1111/febs.13049. PMID 25227262.
- ↑ 9.0 9.1 "The three-dimensional structure of the N-acetylglucosamine-6-phosphate deacetylase, NagA, from Bacillus subtilis: a member of the urease superfamily". The Journal of Biological Chemistry 279 (4): 2809–16. January 2004. doi:10.1074/jbc.M310165200. PMID 14557261.
- ↑ 10.0 10.1 10.2 "How bacteria consume their own exoskeletons (turnover and recycling of cell wall peptidoglycan)". Microbiology and Molecular Biology Reviews 72 (2): 211–27, table of contents. June 2008. doi:10.1128/MMBR.00027-07. PMID 18535144.
- ↑ 11.0 11.1 11.2 "An alternative route for recycling of N-acetylglucosamine from peptidoglycan involves the N-acetylglucosamine phosphotransferase system in Escherichia coli". Journal of Bacteriology 191 (18): 5641–7. September 2009. doi:10.1128/JB.00448-09. PMID 19617367.
- ↑ "Enzymes of UDP-GlcNAc biosynthesis in yeast". Yeast 23 (1): 1–14. January 2006. doi:10.1002/yea.1337. PMID 16408321.
- ↑ 13.0 13.1 "Cell-wall recycling and synthesis in Escherichia coli and Pseudomonas aeruginosa - their role in the development of resistance". Journal of Medical Microbiology 67 (1): 1–21. January 2018. doi:10.1099/jmm.0.000636. PMID 29185941.
- ↑ Stryer, Lubert; Tymoczko, John L.; Berg, Jeremy M. (2002). "The Citric Acid Cycle". Biochemistry. 5th Edition. https://www.ncbi.nlm.nih.gov/books/NBK21163/.
- ↑ 15.0 15.1 "The purification and properties of N-acetylglucosamine 6-phosphate deacetylase from Escherichia coli". The Biochemical Journal 105 (1): 121–5. October 1967. doi:10.1042/bj1050121. PMID 4861885.
- ↑ "Molecular insights of NagA enzyme could help combat TB". 2018-07-12. https://www.news-medical.net/news/20180712/Molecular-insights-of-NagA-enzyme-could-help-combat-TB.aspx.
Further reading
- "Purification and characterization of N-acetylglucosamine 6-phosphate deacetylase with activity against N-acetylglucosamine from Vibrio cholerae non-O1". Bioscience, Biotechnology, and Biochemistry 60 (8): 1320–3. August 1996. doi:10.1271/bbb.60.1320. PMID 8987551.
 |  |

