Earth:Biomineralization
Biomineralization, also written biomineralisation, is the process by which living organisms produce minerals,[lower-alpha 1] often resulting in hardened or stiffened mineralized tissues. It is an extremely widespread phenomenon: all six taxonomic kingdoms contain members that can form minerals, and over 60 different minerals have been identified in organisms.[2][3][4] Examples include silicates in algae and diatoms, carbonates in invertebrates, and calcium phosphates and carbonates in vertebrates. These minerals often form structural features such as sea shells and the bone in mammals and birds.
Organisms have been producing mineralized skeletons for the past 550 million years. Calcium carbonates and calcium phosphates are usually crystalline, but silica organisms (such as sponges and diatoms) are always non-crystalline minerals. Other examples include copper, iron, and gold deposits involving bacteria. Biologically formed minerals often have special uses such as magnetic sensors in magnetotactic bacteria (Fe3O4), gravity-sensing devices (CaCO3, CaSO4, BaSO4) and iron storage and mobilization (Fe2O3•H2O in the protein ferritin).
In terms of taxonomic distribution, the most common biominerals are the phosphate and carbonate salts of calcium that are used in conjunction with organic polymers such as collagen and chitin to give structural support to bones and shells.[5] The structures of these biocomposite materials are highly controlled from the nanometer to the macroscopic level, resulting in complex architectures that provide multifunctional properties. Because this range of control over mineral growth is desirable for materials engineering applications, there is interest in understanding and elucidating the mechanisms of biologically-controlled biomineralization.[6][7]
Types
Mineralization can be subdivided into different categories depending on the following: the organisms or processes that create chemical conditions necessary for mineral formation, the origin of the substrate at the site of mineral precipitation, and the degree of control that the substrate has on crystal morphology, composition, and growth.[8] These subcategories include biomineralization, organomineralization, and inorganic mineralization, which can be subdivided further. However, the usage of these terms varies widely in the scientific literature because there are no standardized definitions. The following definitions are based largely on a paper written by Dupraz et al. (2009),[8] which provided a framework for differentiating these terms.
Biomineralization
Biomineralization, biologically controlled mineralization, occurs when crystal morphology, growth, composition, and location are completely controlled by the cellular processes of a specific organism. Examples include the shells of invertebrates, such as molluscs and brachiopods. Additionally, the mineralization of collagen provides crucial compressive strength for the bones, cartilage, and teeth of vertebrates.[9]
Organomineralization
This type of mineralization includes both biologically induced mineralization and biologically influenced mineralization.
- Biologically induced mineralization occurs when the metabolic activity of microbes (e.g. bacteria) produces chemical conditions favorable for mineral formation. The substrate for mineral growth is the organic matrix, secreted by the microbial community, and affects crystal morphology and composition. Examples of this type of mineralization include calcareous or siliceous stromatolites and other microbial mats. A more specific type of biologically induced mineralization, remote calcification or remote mineralization, takes place when calcifying microbes occupy a shell-secreting organism and alter the chemical environment surrounding the area of shell formation. The result is mineral formation not strongly controlled by the cellular processes of the animal host (i.e., remote mineralization); this may lead to unusual crystal morphologies.[10]
- Biologically influenced mineralization takes place when chemical conditions surrounding the site of mineral formation are influenced by abiotic processes (e.g., evaporation or degassing). However, the organic matrix (secreted by microorganisms) is responsible for crystal morphology and composition. Examples include micro- to nanometer-scale crystals of various morphologies.[11][12]
Biological mineralization can also take place as a result of fossilization. See also Calcification.
Biological roles
Among animals, biominerals composed of calcium carbonate, calcium phosphate, or silica perform a variety of roles such as support, defense, and feeding.[13]
-
 Many protists, like this coccolithophore, have protective mineralised shells.
Many protists, like this coccolithophore, have protective mineralised shells. -
 Forams from a beach
Forams from a beach -
 Many invertebrate animals have external exoskeletons or shells, which achieve rigidity by a variety of mineralisations.
Many invertebrate animals have external exoskeletons or shells, which achieve rigidity by a variety of mineralisations. -
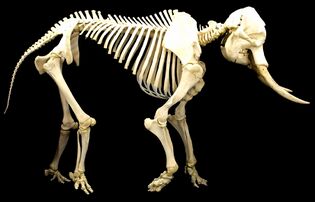 Vertebrate animals have internal endoskeletons which achieve rigidity by binding calcium phosphate into hydroxylapatite.
Vertebrate animals have internal endoskeletons which achieve rigidity by binding calcium phosphate into hydroxylapatite.




If present on a supracellular scale, biominerals are usually deposited by a dedicated organ, which is often defined very early in embryological development. This organ will contain an organic matrix that facilitates and directs the deposition of crystals.[13] The matrix may be collagen, as in deuterostomes,[13] or based on chitin or other polysaccharides, as in molluscs.[14]
In molluscs

The mollusc shell is a biogenic composite material that has been the subject of much interest in materials science because of its unusual properties and its model character for biomineralization. Molluscan shells consist of 95–99% calcium carbonate by weight, while an organic component makes up the remaining 1–5%. The resulting composite has a fracture toughness ≈3000 times greater than that of the crystals themselves.[15] In the biomineralization of the mollusc shell, specialized proteins are responsible for directing crystal nucleation, phase, morphology, and growths dynamics and ultimately give the shell its remarkable mechanical strength. The application of biomimetic principles elucidated from mollusc shell assembly and structure may help in fabricating new composite materials with enhanced optical, electronic, or structural properties. The most described arrangement in mollusc shells is the nacre, known in large shells such as Pinna or the pearl oyster (Pinctada). Not only does the structure of the layers differ, but so do their mineralogy and chemical composition. Both contain organic components (proteins, sugars, and lipids), and the organic components are characteristic of the layer and of the species.[4] The structures and arrangements of mollusc shells are diverse, but they share some features: the main part of the shell is crystalline calcium carbonate (aragonite, calcite), though some amorphous calcium carbonate occurs as well; and although they react as crystals, they never show angles and facets.[16]
In fungi

(b) Fungi as heterotrophs, recycle organic matter. While doing so, they produce metabolites such as organic acids that can also precipitate as secondary minerals (salts). Recycling organic matter eventually releases constitutive elements such as C, N, P, and S
(c) CO2 produced by heterotrophic fungal respiration can dissolve into H2O and depending on the physicochemical conditions precipitate as CaCO3 leading to the formation of a secondary mineral.
Fungi are a diverse group of organisms that belong to the eukaryotic domain. Studies of their significant roles in geological processes, "geomycology", have shown that fungi are involved with biomineralization, biodegradation, and metal-fungal interactions.[18]
In studying fungi's roles in biomineralization, it has been found that fungi deposit minerals with the help of an organic matrix, such as a protein, that provides a nucleation site for the growth of biominerals.[19] Fungal growth may produce a copper-containing mineral precipitate, such as copper carbonate produced from a mixture of (NH4)2CO3 and CuCl2.[19] The production of the copper carbonate is produced in the presence of proteins made and secreted by the fungi.[19] These fungal proteins that are found extracellularly aid in the size and morphology of the carbonate minerals precipitated by the fungi.[19]
In addition to precipitating carbonate minerals, fungi can also precipitate uranium-containing phosphate biominerals in the presence of organic phosphorus that acts as a substrate for the process.[20] The fungi produce a hyphal matrix, also known as mycelium, that localizes and accumulates the uranium minerals that have been precipitated.[20] Although uranium is often deemed toxic to living organisms, certain fungi such as Aspergillus niger and Paecilomyces javanicus can tolerate it.[20]
Though minerals can be produced by fungi, they can also be degraded, mainly by oxalic acid–producing strains of fungi.[21] Oxalic acid production is increased in the presence of glucose for three organic acid producing fungi: Aspergillus niger, Serpula himantioides, and Trametes versicolor.[21] These fungi have been found to corrode apatite and galena minerals.[21] Degradation of minerals by fungi is carried out through a process known as neogenesis.[22] The order of most to least oxalic acid secreted by the fungi studied are Aspergillus niger, followed by Serpula himantioides, and finally Trametes versicolor.[21]
In bacteria
It is less clear what purpose biominerals serve in bacteria. One hypothesis is that cells create them to avoid entombment by their own metabolic byproducts. Iron oxide particles may also enhance their metabolism.[23]
Other roles

Biomineralization plays significant global roles terraforming the planet, as well as in biogeochemical cycles[17] and as a carbon sink.[24]
Composition
Most biominerals can be grouped by chemical composition into one of three distinct mineral classes: silicates, carbonates, or phosphates.[25]
Silicates
File:Collection Penard MHNG Specimen 533-2-1 Pamphagus granulatus.tif

Silicates (glass) are common in marine biominerals, where diatoms form frustules and radiolaria form capsules from hydrated amorphous silica (opal).[27]
Carbonates
The major carbonate in biominerals is CaCO3. The most common polymorphs in biomineralization are calcite (e.g. foraminifera, coccolithophores) and aragonite (e.g. corals), although metastable vaterite and amorphous calcium carbonate can also be important, either structurally[28][29] or as intermediate phases in biomineralization.[30][31] Some biominerals include a mixture of these phases in distinct, organised structural components (e.g. bivalve shells). Carbonates are particularly prevalent in marine environments, but also present in freshwater and terrestrial organisms.[32]
Phosphates
The most common biogenic phosphate is hydroxyapatite (HA), a calcium phosphate (Ca10(PO4)6(OH)2) and a naturally occurring form of apatite. It is a primary constituent of bone, teeth, and fish scales.[33] Bone is made primarily of HA crystals interspersed in a collagen matrix—65 to 70% of the mass of bone is HA. Similarly, HA is 70 to 80% of the mass of dentin and enamel in teeth. In enamel, the matrix for HA is formed by amelogenins and enamelins instead of collagen.[34] Remineralisation of tooth enamel involves the reintroduction of mineral ions into demineralised enamel.[35] Hydroxyapatite is the main mineral component of enamel in teeth.[36] During demineralisation, calcium and phosphorus ions are drawn out from the hydroxyapatite. The mineral ions introduced during remineralisation restore the structure of the hydroxyapatite crystals.[36]
The clubbing appendages of the peacock mantis shrimp are made of an extremely dense form of the mineral which has a higher specific strength; this has led to its investigation for potential synthesis and engineering use.[37] Their dactyl appendages have excellent impact resistance due to the impact region being composed of mainly crystalline hydroxyapatite, which offers significant hardness. A periodic layer underneath the impact layer composed of hydroxyapatite with lower calcium and phosphorus content (thus resulting in a much lower modulus) inhibits crack growth by forcing new cracks to change directions. This periodic layer also reduces the energy transferred across both layers due to the large difference in modulus, even reflecting some of the incident energy.[38]

| Composition | Example organisms |
|---|---|
| Calcium carbonate (calcite or aragonite) |
|
| Silica (silicate/glass/opal) |
|
| Apatite (phosphate minerals) |
|
Other minerals
Beyond these main three categories, there are a number of less-common types of biominerals, usually resulting from a need for specific physical properties or the organism inhabiting an unusual environment. For example, teeth that are primarily used for scraping hard substrates may be reinforced with particularly tough minerals, such as the iron minerals magnetite in chitons[39] or goethite in limpets.[40] Gastropod molluscs living close to hydrothermal vents reinforce their carbonate shells with the iron-sulfur minerals pyrite and greigite.[41] Magnetotactic bacteria also employ magnetic iron minerals magnetite and greigite to produce magnetosomes to aid orientation and distribution in the sediments.[42][43]

-
![Chitons have aragonite shells and aragonite-based eyes,[45] as well as teeth coated with magnetite.](/wiki/images/thumb/3/39/Chitonidae_-_Chiton_squamosus.JPG/280px-Chitonidae_-_Chiton_squamosus.JPG)
-
 Limpets have carbonate shells and teeth reinforced with goethite.
Limpets have carbonate shells and teeth reinforced with goethite.
![Chitons have aragonite shells and aragonite-based eyes,[45] as well as teeth coated with magnetite.](/wiki/File:Chitonidae_-_Chiton_squamosus.JPG)

-
 Acantharian radiolarians have celestine crystal shells.
Acantharian radiolarians have celestine crystal shells. -
 Celestine crystals, the heaviest mineral in the oceans
Celestine crystals, the heaviest mineral in the oceans


Celestine, the heaviest mineral in the ocean, consists of strontium sulfate, SrSO4. The mineral is named for the delicate blue colour of its crystals.[46] Planktic acantharean radiolarians form celestine crystal shells. The denseness of the celestite ensures their shells function as mineral ballast, resulting in fast sedimentation to bathypelagic depths. High settling fluxes of acantharian cysts have been observed at times in the Iceland Basin and the Southern Ocean, as much as half of the total gravitational organic carbon flux.[47][48][46]
Diversity


_mitra_Ehrenberg_-_160x.jpg)
In nature, there is a wide array of biominerals, ranging from iron oxide to strontium sulfate,[49] with calcareous biominerals being particularly notable.[50][51] However, the most taxonomically widespread biomineral is silica (SiO2·nH2O), being present in all eukaryotic supergroups.[52] Notwithstanding, the degree of silicification can vary even between closely related taxa, from being found in composite structures with other biominerals (e.g., limpet teeth;[53] to forming minor structures (e.g., ciliate granules;[54] or being a major structural constituent of the organism.[55] The most extreme degree of silicification is evident in the diatoms, where almost all species have an obligate requirement for silicon to complete cell wall formation and cell division.[56][57] Biogeochemically and ecologically, diatoms are the most important silicifiers in modern marine ecosystems, with radiolarians (polycystine and phaeodarian rhizarians), silicoflagellates (dictyochophyte and chrysophyte stramenopiles), and sponges with prominent roles as well. In contrast, the major silicifiers in terrestrial ecosystems are the land plants (embryophytes), with other silicifying groups (e.g., testate amoebae) having a minor role.[58]
Broadly, biomineralized structures evolve and diversify when the energetic cost of biomineral production is less than the expense of producing an equivalent organic structure.[59][60][61] The energetic costs of forming a silica structure from silicic acid are much less than forming the same volume from an organic structure (≈20-fold less than lignin or 10-fold less than polysaccharides like cellulose).[62] Based on a structural model of biogenic silica,[63] Lobel et al. (1996) identified by biochemical modeling a low-energy reaction pathway for nucleation and growth of silica.[64] The combination of organic and inorganic components within biomineralized structures often results in enhanced properties compared to exclusively organic or inorganic materials. With respect to biogenic silica, this can result in the production of much stronger structures, such as siliceous diatom frustules having the highest strength per unit density of any known biological material,[65][66] or sponge spicules being many times more flexible than an equivalent structure made of pure silica.[67][68] As a result, biogenic silica structures are used for support,[69] feeding,[70] predation defense[71][72][73] and environmental protection as a component of cyst walls.[55] Biogenic silica also has useful optical properties for light transmission and modulation in organisms as diverse as plants,[74] diatoms,[75][76][77] sponges,[78] and molluscs.[79] There is also evidence that silicification is used as a detoxification response in snails[80] and plants,[81] biosilica has even been suggested to play a role as a pH buffer for the enzymatic activity of carbonic anhydrase, aiding the acquisition of inorganic carbon for photosynthesis.[82][58]
-
![Diversity of biomineralization across the eukaryotes[58] The phylogeny shown in this diagram is based on Adl et al. (2012),[83] with major eukaryotic supergroups named in boxes. Letters next to taxon names denote the presence of biomineralization, with circled letters indicating the prominent and widespread use of that biomineral. S, silica; C, calcium carbonate; P, calcium phosphate; I, iron (magnetite/goethite); X, calcium oxalate; SO4, sulfates (calcium/barium/strontium), ? denotes uncertainty in the report.[84][85][25][52][49][86]](/wiki/images/0/0f/Diversity_of_biomineralization_across_the_eukaryotes.jpg) Diversity of biomineralization across the eukaryotes[58]The phylogeny shown in this diagram is based on Adl et al. (2012),[83] with major eukaryotic supergroups named in boxes. Letters next to taxon names denote the presence of biomineralization, with circled letters indicating the prominent and widespread use of that biomineral. S, silica; C, calcium carbonate; P, calcium phosphate; I, iron (magnetite/goethite); X, calcium oxalate; SO4, sulfates (calcium/barium/strontium), ? denotes uncertainty in the report.[84][85][25][52][49][86]
Diversity of biomineralization across the eukaryotes[58]The phylogeny shown in this diagram is based on Adl et al. (2012),[83] with major eukaryotic supergroups named in boxes. Letters next to taxon names denote the presence of biomineralization, with circled letters indicating the prominent and widespread use of that biomineral. S, silica; C, calcium carbonate; P, calcium phosphate; I, iron (magnetite/goethite); X, calcium oxalate; SO4, sulfates (calcium/barium/strontium), ? denotes uncertainty in the report.[84][85][25][52][49][86]
![Diversity of biomineralization across the eukaryotes[58] The phylogeny shown in this diagram is based on Adl et al. (2012),[83] with major eukaryotic supergroups named in boxes. Letters next to taxon names denote the presence of biomineralization, with circled letters indicating the prominent and widespread use of that biomineral. S, silica; C, calcium carbonate; P, calcium phosphate; I, iron (magnetite/goethite); X, calcium oxalate; SO4, sulfates (calcium/barium/strontium), ? denotes uncertainty in the report.[84][85][25][52][49][86]](/wiki/File:Diversity_of_biomineralization_across_the_eukaryotes.jpg)
There are questions which have yet to be resolved, such as why some organisms biomineralize while others do not, and why is there such a diversity of biominerals besides silicon when silicon is so abundant, comprising 28% of the Earth's crust.[58] The answer to these questions lies in the evolutionary interplay between biomineralization and geochemistry, and in the competitive interactions that have arisen from these dynamics. Fundamentally whether an organism produces silica or not involves evolutionary trade-offs and competition between silicifiers themselves, and non-silicifying organisms (both those which use other biominerals, and non-mineralizing groups). Mathematical models and controlled experiments of resource competition in phytoplankton have demonstrated the rise to dominance of different algal species based on nutrient backgrounds in defined media. These have been part of fundamental studies in ecology.[87][88] However, the vast diversity of organisms that thrive in a complex array of biotic and abiotic interactions in oceanic ecosystems are a challenge to such minimal models and experimental designs, whose parameterization and possible combinations, respectively, limit the interpretations that can be built on them.[58]
Evolution

The first evidence of biomineralization dates to some 750 million years ago,[89][90] and sponge-grade organisms may have formed calcite skeletons 630 million years ago.[91] But in most lineages, biomineralization first occurred in the Cambrian or Ordovician periods.[92] Organisms used whichever form of calcium carbonate was more stable in the water column at the point in time when they became biomineralized,[93] and stuck with that form for the remainder of their biological history[94] (but see[95] for a more detailed analysis). The stability is dependent on the Ca/Mg ratio of seawater, which is thought to be controlled primarily by the rate of sea floor spreading, although atmospheric CO
2 levels may also play a role.[93]
Biomineralization evolved multiple times, independently,[96] and most animal lineages first expressed biomineralized components in the Cambrian period.[97] Many of the same processes are used in unrelated lineages, which suggests that biomineralization machinery was assembled from pre-existing "off-the-shelf" components already used for other purposes in the organism.[25] Although the biomachinery facilitating biomineralization is complex – involving signalling transmitters, inhibitors, and transcription factors – many elements of this 'toolkit' are shared between phyla as diverse as corals, molluscs, and vertebrates.[98] The shared components tend to perform quite fundamental tasks, such as designating that cells will be used to create the minerals, whereas genes controlling more finely tuned aspects that occur later in the biomineralization process, such as the precise alignment and structure of the crystals produced, tend to be uniquely evolved in different lineages.[13][99] This suggests that Precambrian organisms were employing the same elements, albeit for a different purpose – perhaps to avoid the inadvertent precipitation of calcium carbonate from the supersaturated Proterozoic oceans.[98] Forms of mucus that are involved in inducing mineralization in most animal lineages appear to have performed such an anticalcifatory function in the ancestral state.[100] Further, certain proteins that would originally have been involved in maintaining calcium concentrations within cells[101] are homologous in all animals, and appear to have been co-opted into biomineralization after the divergence of the animal lineages.[102] The galaxins are one probable example of a gene being co-opted from a different ancestral purpose into controlling biomineralization, in this case, being 'switched' to this purpose in the Triassic scleractinian corals; the role performed appears to be functionally identical to that of the unrelated pearlin gene in molluscs.[103] Carbonic anhydrase serves a role in mineralization broadly in the animal kingdom, including in sponges, implying an ancestral role.[104] Far from being a rare trait that evolved a few times and remained stagnant, biomineralization pathways in fact evolved many times and are still evolving rapidly today; even within a single genus, it is possible to detect great variation within a single gene family.[99]
_2.jpg)
The homology of biomineralization pathways is underlined by a remarkable experiment whereby the nacreous layer of a molluscan shell was implanted into a human tooth, and rather than experiencing an immune response, the molluscan nacre was incorporated into the host bone matrix. This points to the exaptation of an original biomineralization pathway. The biomineralisation capacity of brachiopods and molluscs has also been demonstrated to be homologous, building on a conserved set of genes.[105]
The most ancient example of biomineralization, dating back 2 billion years, is the deposition of magnetite, which is observed in some bacteria, as well as the teeth of chitons and the brains of vertebrates; it is possible that this pathway, which performed a magnetosensory role in the common ancestor of all bilaterians, was duplicated and modified in the Cambrian to form the basis for calcium-based biomineralization pathways.[106] Iron is stored in close proximity to magnetite-coated chiton teeth, so that the teeth can be renewed as they wear. Not only is there a marked similarity between the magnetite deposition process and enamel deposition in vertebrates, but some vertebrates even have comparable iron storage facilities near their teeth.[107]
Potential applications
Most traditional approaches to the synthesis of nanoscale materials are energy inefficient, requiring stringent conditions (e.g., high temperature, pressure, or pH), and often produce toxic byproducts. Furthermore, the quantities produced are small, and the resultant material is usually irreproducible because of the difficulties in controlling agglomeration.[108] In contrast, materials produced by organisms have properties that usually surpass those of analogous synthetically manufactured materials with similar phase composition. Biological materials are assembled in aqueous environments under mild conditions by using macromolecules. Organic macromolecules collect and transport raw materials and assemble these substrates and into short- and long-range ordered composites with consistency and uniformity.[109][110]
The aim of biomimetics is to mimic the natural way of producing minerals such as apatites. Many man-made crystals require elevated temperatures and strong chemical solutions, whereas the organisms have long been able to lay down elaborate mineral structures at ambient temperatures. Often, the mineral phases are not pure but are made as composites that entail an organic part, often protein, which takes part in and controls the biomineralization. These composites are often not only as hard as the pure mineral but also tougher, as the micro-environment controls biomineralization.[109][110]
Architecture


One biological system that might be of key importance in the future development of architecture is bacterial biofilm. The term biofilm refers to complex heterogeneous structures comprising different populations of microorganisms that attach and form a community on inert (e.g. rocks, glass, plastic) or organic (e.g. skin, cuticle, mucosa) surfaces.[111]
The properties of the surface, such as charge, hydrophobicity, and roughness, determine initial bacterial attachment.[112] A common principle of all biofilms is the production of extracellular matrix (ECM) composed of different organic substances, such as extracellular proteins, exopolysaccharides, and nucleic acids.[113] While the ability to generate ECM appears to be a common feature of multicellular bacterial communities, the means by which these matrices are constructed and function are diverse.[113][114][115][116]
-
![Model for biomineralization-mediated scaffolding of bacterial biofilms A directed growth of the calcium carbonate crystals allows mechanical support of the 3D structure. The bacterial extracellular matrix (brown) promotes the crystals' growth in specific directions.[117][116]](/wiki/images/thumb/e/ee/Biomineralization%E2%80%90mediated_scaffolding_of_bacterial_biofilms.jpg/576px-Biomineralization%E2%80%90mediated_scaffolding_of_bacterial_biofilms.jpg) Model for biomineralization-mediated scaffoldingA directed growth of the calcium carbonate crystals allows mechanical support of the 3D structure. The bacterial extracellular matrix (brown) promotes the crystals' growth in specific directions.[117][116]
Model for biomineralization-mediated scaffoldingA directed growth of the calcium carbonate crystals allows mechanical support of the 3D structure. The bacterial extracellular matrix (brown) promotes the crystals' growth in specific directions.[117][116]
of bacterial biofilms
![Model for biomineralization-mediated scaffolding of bacterial biofilms A directed growth of the calcium carbonate crystals allows mechanical support of the 3D structure. The bacterial extracellular matrix (brown) promotes the crystals' growth in specific directions.[117][116]](/wiki/File:Biomineralization%E2%80%90mediated_scaffolding_of_bacterial_biofilms.jpg)
Bacterially induced calcium carbonate precipitation can be used to produce "self-healing" concrete. Bacillus megaterium spores and suitable dried nutrients are mixed and applied to steel-reinforced concrete. When the concrete cracks, water ingress dissolves the nutrients and the bacteria germinate triggering calcium carbonate precipitation, resealing the crack and protecting the steel reinforcement from corrosion.[118] This process can also be used to manufacture new hard materials, such as bio-cement.[119][116]
However, the full potential of bacteria-driven biomineralization is yet to be realized, as it is currently used as a passive filling rather than as a smart designable material. A future challenge is to develop ways to control the timing and the location of mineral formation, as well as the physical properties of the mineral itself, by environmental input. Bacillus subtilis has already been shown to respond to its environment, by changing the production of its ECM. It uses the polymers produced by single cells during biofilm formation as a physical cue to coordinate ECM production by the bacterial community.[120][121][116]
Uranium contaminants
-
 Autunite crystal
Autunite crystal

Biomineralization may be used to remediate groundwater contaminated with uranium.[122] The biomineralization of uranium primarily involves the precipitation of uranium phosphate minerals associated with the release of phosphate by microorganisms. Negatively charged ligands at the surface of the cells attract the positively charged uranyl ion (UO22+). If the concentrations of phosphate and UO22+ are sufficiently high, minerals such as autunite (Ca(UO2)2(PO4)2•10-12H2O) or polycrystalline HUO2PO4 may form thus reducing the mobility of UO22+. Compared to the direct addition of inorganic phosphate to contaminated groundwater, biomineralization has the advantage that the ligands produced by microbes will target uranium compounds more specifically rather than react actively with all aqueous metals. Stimulating bacterial phosphatase activity to liberate phosphate under controlled conditions limits the rate of bacterial hydrolysis of organophosphate and the release of phosphate to the system, thus avoiding clogging of the injection location with metal phosphate minerals.[122] The high concentration of ligands near the cell surface also provides nucleation foci for precipitation, which leads to higher efficiency than chemical precipitation.[123]
Biogenic mineral controversy
The International Mineralogical Association (IMA) is the generally recognized standard body for the definition and nomenclature of mineral species. As of December 2020[update], the IMA recognizes 5,650 official mineral species[124] out of 5,862 proposed or traditional ones.[125]
The IMA's decision to exclude biogenic crystalline substances is a topic of contention among geologists and mineralogists. For example, Lowenstam (1981) stated that "organisms are capable of forming a diverse array of minerals, some of which cannot be formed inorganically in the biosphere."[126]
Skinner (2005) views all solids as potential minerals and includes biominerals in the mineral kingdom, which are created by organisms' metabolic activities. Skinner expanded the previous definition of a mineral to classify "element or compound, amorphous or crystalline, formed through biogeochemical processes," as a mineral.[127]
Recent advances in high-resolution genetics and X-ray absorption spectroscopy are providing revelations on the biogeochemical relations between microorganisms and minerals that may shed new light on this question.[128][127] For example, the IMA-commissioned "Working Group on Environmental Mineralogy and Geochemistry " deals with minerals in the hydrosphere, atmosphere, and biosphere.[129] The group's scope includes mineral-forming microorganisms, which exist on nearly every rock, soil, and particle surface spanning the globe to depths of at least 1,600 metres below the sea floor and 70 kilometres into the stratosphere (possibly entering the mesosphere).[130][131][132]
Biogeochemical cycles have contributed to the formation of minerals for billions of years. Microorganisms can precipitate metals from solution, contributing to the formation of ore deposits. They can also catalyze the dissolution of minerals.[133][134][135]
Before the International Mineralogical Association's listing, over 60 biominerals had been discovered, named, and published.[136] These minerals (a sub-set tabulated in Lowenstam (1981)[126]) are considered minerals proper according to Skinner's (2005) definition.[127] These biominerals are not listed in the International Mineral Association official list of mineral names,[137] however, many of these biomineral representatives are distributed among the 78 mineral classes listed in the Dana classification scheme.[127]
Skinner's (2005) definition of a mineral considers this matter by stating that a mineral can be crystalline or amorphous.[127] Although biominerals are not the most common form of minerals,[138] they help to define the limits of what constitutes a mineral properly. Nickel's (1995) formal definition explicitly mentioned crystallinity as a key to defining a substance as a mineral.[128] A 2011 article defined icosahedrite, an aluminium-iron-copper alloy as mineral; named for its unique natural icosahedral symmetry, it is a quasicrystal. Unlike a true crystal, quasicrystals are ordered but not periodic.[139][140]
List of minerals
Examples of biogenic minerals include:[141]
- Apatite in bones and teeth
- Aragonite, calcite, fluorite in vestibular systems (part of the inner ear) of vertebrates
- Aragonite and calcite in travertine and biogenic silica (siliceous sinter, opal) deposited through algal action
- Goethite found as filaments in limpet teeth
- Hydroxyapatite formed by mitochondria
- Magnetite and greigite formed by magnetotactic bacteria
- Oxalate and calcium carbonate raphides, silica bodies, strontium and barium sulfate in some plants[142]
- Pyrite and marcasite in sedimentary rocks deposited by sulfate-reducing bacteria
- Quartz formed from bacterial action on fossil fuels (gas, oil, coal)
Astrobiology
Biominerals could be important indicators of extraterrestrial life and thus could play an essential role in the search for past or present life on Mars. Furthermore, organic components (biosignatures) that are often associated with biominerals are believed to play crucial roles in both pre-biotic and biotic reactions.[143]
On 24 January 2014, NASA reported that current studies by the Curiosity and Opportunity rovers on the planet Mars will now be searching for evidence of ancient life, including a biosphere based on autotrophic, chemotrophic and chemolithoautotrophic microorganisms, as well as ancient water, including fluvio-lacustrine environments (plains related to ancient rivers or lakes) that may have been habitable.[144][145][146][147] The search for evidence of habitability, taphonomy (related to fossils), and organic carbon on the planet Mars is now a primary NASA objective.[144][145]
See also
- Biocrystallization
- Biofilm
- Biointerface
- Biomineralising polychaetes
- Bone mineral
- Microbiologically induced calcite precipitation
- Micropaleontology
- Mineralized tissues
- Phytolith
- Raphide and Druse (botany)
- Susannah M. Porter history of biomineralization
- Malcolm L. Snead Researcher on amelogenin and dental enamel biomineralization
Notes
- ↑ The International Union of Pure and Applied Chemistry defines biomineralization as "mineralization caused by cell-mediated phenomena" and notes that it "is a process generally concomitant to biodegradation".[1]
References
- ↑ Cite error: Invalid
<ref>tag; no text was provided for refs namedVert_2012 - ↑ Biomineralization: From Nature to Application. Metal Ions in Life Sciences. 4. Wiley. 2008. ISBN 978-0-470-03525-2.
- ↑ On biomineralization. Oxford [Oxfordshire]: Oxford University Press. 1989. ISBN 978-0-19-504977-0.
- ↑ 4.0 4.1 Biominerals and fossils through time. Cambridge. 2011. ISBN 978-0-521-87473-1.
- ↑ "Occurrence, formation and function of organic sheets in the mineral tube structures of Serpulidae (polychaeta, Annelida)". PLOS ONE 8 (10). 2013. doi:10.1371/journal.pone.0075330. PMID 24116035. Bibcode: 2013PLoSO...875330V.
- ↑ "Biomineralization: conflicts, challenges, and opportunities". Journal of Cellular Biochemistry 30-31 (S30-31): 83–91. 1998. doi:10.1002/(SICI)1097-4644(1998)72:30/31+<83::AID-JCB12>3.0.CO;2-F. PMID 9893259.
- ↑ "Biomimetics: materials fabrication through biology". Proceedings of the National Academy of Sciences of the United States of America 96 (25): 14183–14185. December 1999. doi:10.1073/pnas.96.25.14183. PMID 10588672. Bibcode: 1999PNAS...9614183S.
- ↑ 8.0 8.1 "Processes of carbonate precipitation in modern microbial mats". Earth-Science Reviews. Microbial Mats in Earth's Fossil Record of Life: Geobiology 96 (3): 141–162. 1 October 2009. doi:10.1016/j.earscirev.2008.10.005. Bibcode: 2009ESRv...96..141D.
- ↑ "The materials science of collagen". Journal of the Mechanical Behavior of Biomedical Materials 52: 22–50. December 2015. doi:10.1016/j.jmbbm.2015.05.023. PMID 26144973.
- ↑ "The oyster enigma variations: a hypothesis of microbial calcification". Paleobiology 40 (1): 1–13. 27 September 2013. doi:10.1666/13002. ISSN 0094-8373. https://escholarship.org/uc/item/8kn6h5dg.
- ↑ Bindschedler, Saskia; Cailleau, Guillaume; Verrecchia, Eric (2016-05-05). "Role of Fungi in the Biomineralization of Calcite". Minerals (MDPI AG) 6 (2): 41. doi:10.3390/min6020041. ISSN 2075-163X. Bibcode: 2016Mine....6...41B.
- ↑ Görgen, Sigrid; Benzerara, Karim; Skouri-Panet, Fériel; Gugger, Muriel; Chauvat, Franck; Cassier-Chauvat, Corinne (2020-11-30). "The diversity of molecular mechanisms of carbonate biomineralization by bacteria". Discover Materials (Springer Science and Business Media LLC) 1 (1). doi:10.1007/s43939-020-00001-9. ISSN 2730-7727.
- ↑ 13.0 13.1 13.2 13.3 "A genome-wide analysis of biomineralization-related proteins in the sea urchin Strongylocentrotus purpuratus". Developmental Biology 300 (1): 335–348. December 2006. doi:10.1016/j.ydbio.2006.07.047. PMID 16987510.
- ↑ "Nacre and false nacre (foliated aragonite) in extant monoplacophorans (=Tryblidiida: Mollusca)". Die Naturwissenschaften 96 (1): 111–122. January 2009. doi:10.1007/s00114-008-0461-1. PMID 18843476. Bibcode: 2009NW.....96..111C.
- ↑ "The design of mineralised hard tissues for their mechanical functions". The Journal of Experimental Biology 202 (Pt 23): 3285–3294. December 1999. doi:10.1242/jeb.202.23.3285. PMID 10562511. Bibcode: 1999JExpB.202.3285C.
- ↑ Les étapes de la découverte des rapports entre la terre et la vie: une introduction à la paléontologie. Paris: Éditions scientifiques GB. 2003. ISBN 978-2-84703-008-2. OCLC 77036366.
- ↑ 17.0 17.1 "Programming pluripotent precursor cells derived from Xenopus embryos to generate specific tissues and organs". Genes (MDPI AG) 1 (3): 413–426. November 2010. doi:10.3390/min6020041. PMID 24710095. Bibcode: 2016Mine....6...41B. 50px Modified material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ "Geomycology: biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation". Mycological Research 111 (Pt 1): 3–49. January 2007. doi:10.1016/j.mycres.2006.12.001. PMID 17307120.
- ↑ 19.0 19.1 19.2 19.3 "Biosynthesis of copper carbonate nanoparticles by ureolytic fungi". Applied Microbiology and Biotechnology 101 (19): 7397–7407. October 2017. doi:10.1007/s00253-017-8451-x. PMID 28799032.
- ↑ 20.0 20.1 20.2 "Uranium phosphate biomineralization by fungi". Environmental Microbiology 17 (6): 2064–2075. June 2015. doi:10.1111/1462-2920.12771. PMID 25580878. Bibcode: 2015EnvMi..17.2064L.
- ↑ 21.0 21.1 21.2 21.3 "Fungal degradation of calcium-, lead- and silicon-bearing minerals". Biometals 18 (3): 269–281. June 2005. doi:10.1007/s10534-005-1539-2. PMID 15984571.
- ↑ "Weathering of rocks and neogenesis of minerals associated with lichen activity". Applied Clay Science 16 (5): 229–256. 1 May 2000. doi:10.1016/S0169-1317(99)00056-3. Bibcode: 2000ApCS...16..229A.
- ↑ "Geochemistry. What biogenic minerals tell us". Science 303 (5664): 1618–1619. March 2004. doi:10.1126/science.1095177. PMID 15016984.
- ↑ "Carbon Sequestration via Biomineralization: Processes, Applications and Future Directions". Sustainable Agriculture Reviews 37. 37. Cham: Springer International Publishing. 2019. pp. 93–106. doi:10.1007/978-3-030-29298-0_5. ISBN 978-3-030-29297-3.
- ↑ 25.0 25.1 25.2 Knoll, A.H. (2004). "Biomineralization and evolutionary history". Reviews in Mineralogy and Geochemistry. http://www.geochem.geos.vt.edu/bgep/pubs/Chapter_11_Knoll.pdf.
- ↑ "Extreme impact and cavitation forces of a biological hammer: strike forces of the peacock mantis shrimp Odontodactylus scyllarus". The Journal of Experimental Biology 208 (Pt 19): 3655–3664. October 2005. doi:10.1242/jeb.01831. PMID 16169943. Bibcode: 2005JExpB.208.3655P.
- ↑ "Marine Silica Cycle". Encyclopedia of Ocean Sciences. 2001. pp. 1659–1667. doi:10.1006/rwos.2001.0278. ISBN 978-0-12-227430-5.
- ↑ "Narrowly Distributed Crystal Orientation in Biomineral Vaterite" (in en). Chemistry of Materials 27 (19): 6516–6523. 13 October 2015. doi:10.1021/acs.chemmater.5b01542. ISSN 0897-4756.
- ↑ "Amorphous and crystalline calcium carbonate distribution in the tergite cuticle of moulting Porcellio scaber (Isopoda, Crustacea)". Journal of Structural Biology 175 (1): 10–20. July 2011. doi:10.1016/j.jsb.2011.03.019. PMID 21458575. https://bib-pubdb1.desy.de/record/140148/files/Amorphous.pdf.
- ↑ "Planktic foraminifera form their shells via metastable carbonate phases". Nature Communications 8 (1): 1265. November 2017. doi:10.1038/s41467-017-00955-0. PMID 29097678. Bibcode: 2017NatCo...8.1265J.
- ↑ "Amorphous calcium carbonate particles form coral skeletons". Proceedings of the National Academy of Sciences of the United States of America 114 (37): E7670–E7678. September 2017. doi:10.1073/pnas.1707890114. PMID 28847944. Bibcode: 2017PNAS..114E7670M.
- ↑ "Biomineralization by photosynthetic organisms: evidence of coevolution of the organisms and their environment?". Geobiology 7 (2): 140–154. March 2009. doi:10.1111/j.1472-4669.2008.00181.x. PMID 19207569. Bibcode: 2009Gbio....7..140R.
- ↑ "Studies on fish scale formation and resorption. III. Fine structure and calcification of the fibrillary plates of the scales in Carassius auratus (Cypriniformes: Cyprinidae)". Cell and Tissue Research 201 (3): 409–422. October 1979. doi:10.1007/BF00236999. PMID 574424.
- ↑ "Biomaterials, Hydroxyapatite". Stat Pearls. January 2018. https://www.ncbi.nlm.nih.gov/books/NBK513314/. Retrieved 12 August 2018.
- ↑ "Demineralization-remineralization dynamics in teeth and bone". International Journal of Nanomedicine 11: 4743–4763. 2016. doi:10.2147/IJN.S107624. PMID 27695330.
- ↑ 36.0 36.1 "Nano-hydroxyapatite and its applications in preventive, restorative and regenerative dentistry: a review of literature". Annali di Stomatologia 5 (3): 108–114. July 2014. PMID 25506416.
- ↑ "The stomatopod dactyl club: a formidable damage-tolerant biological hammer". Science 336 (6086): 1275–1280. June 2012. doi:10.1126/science.1218764. PMID 22679090. Bibcode: 2012Sci...336.1275W. https://www.researchgate.net/publication/225280684. Retrieved 2 December 2017.
- ↑ "Materials science. Small but extremely tough". Science 336 (6086): 1237–1238. June 2012. doi:10.1126/science.1222642. PMID 22679085. Bibcode: 2012Sci...336.1237T.
- ↑ "The Chiton Radula: A Model System for Versatile Use of Iron Oxides*" (in en). Iron Oxides (1st ed.). Wiley. 5 July 2016. pp. 177–206. doi:10.1002/9783527691395.ch8. ISBN 978-3-527-33882-5.
- ↑ "Extreme strength observed in limpet teeth". Journal of the Royal Society, Interface 12 (105). April 2015. doi:10.1098/rsif.2014.1326. PMID 25694539.
- ↑ "The 'scaly-foot gastropod': a new genus and species of hydrothermal vent-endemic gastropod (Neomphalina: Peltospiridae) from the Indian Ocean" (in en). Journal of Molluscan Studies 81 (3): 322–334. August 2015. doi:10.1093/mollus/eyv013. ISSN 0260-1230.
- ↑ Lefèvre, Christopher T.; Bazylinski, Dennis A. (2013). "Ecology, Diversity, and Evolution of Magnetotactic Bacteria". Microbiology and Molecular Biology Reviews 77 (3): 497–526. doi:10.1128/MMBR.00021-13. ISSN 1092-2172. https://journals.asm.org/doi/10.1128/MMBR.00021-13. Retrieved 2026-01-30.
- ↑ Goswami, Pranami; He, Kuang; Li, Jinhua; Pan, Yongxin; Roberts, Andrew P.; Lin, Wei (2022-06-01). "Magnetotactic bacteria and magnetofossils: ecology, evolution and environmental implications". npj Biofilms and Microbiomes (Springer Science and Business Media LLC) 8 (1). doi:10.1038/s41522-022-00304-0. ISSN 2055-5008.
- ↑ "Phylogenetic significance of composition and crystal morphology of magnetosome minerals". Frontiers in Microbiology 4: 344. November 2013. doi:10.3389/fmicb.2013.00344. PMID 24324461.
- ↑ "Weird Sea Mollusk Sports Hundreds of Eyes Made of Armor". 19 November 2015. https://www.livescience.com/52857-mollusk-has-eyes-made-of-armor.html.
- ↑ 46.0 46.1 "Pathways of Organic Carbon Downward Transport by the Oceanic Biological Carbon Pump". Frontiers in Marine Science 6: 634. 2019. doi:10.3389/fmars.2019.00634. Bibcode: 2019FrMaS...6..634L. 50px Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ "Sedimentation of acantharian cysts in the Iceland Basin: Strontium as a ballast for deep ocean particle flux, and implications for acantharian reproductive strategies". Limnology and Oceanography 55 (2): 604–614. 2010. doi:10.4319/lo.2009.55.2.0604.
- ↑ "Acantharian cysts: High flux occurrence in the bathypelagic zone of the Scotia Sea, Southern Ocean". Marine Biology 165 (7): 117. 2018. doi:10.1007/s00227-018-3376-1. Bibcode: 2018MarBi.165..117B. https://nora.nerc.ac.uk/id/eprint/520421/2/Supplementary%20material.pdf.
- ↑ 49.0 49.1 "Non-Skeletal Biomineralization by Eukaryotes: Matters of Moment and Gravity". Geomicrobiology Journal 27 (6–7): 572–584. 2010. doi:10.1080/01490451003702990. Bibcode: 2010GmbJ...27..572R. https://dash.harvard.edu/handle/1/4795339.
- ↑ "Biomineralization and Evolutionary History". Reviews in Mineralogy and Geochemistry 54 (1): 329–356. 2003. doi:10.2113/0540329. Bibcode: 2003RvMG...54..329K.
- ↑ "Protistan Skeletons: A Geologic History of Evolution and Constraint". Evolution of Lightweight Structures. Biologically-Inspired Systems. 6. Dordrecht: Springer. 2015. pp. 1–16. doi:10.1007/978-94-017-9398-8_1. ISBN 978-94-017-9397-1. https://dash.harvard.edu/handle/1/30403697.
- ↑ 52.0 52.1 "The Evolution of Silicon Transport in Eukaryotes". Molecular Biology and Evolution 33 (12): 3226–3248. December 2016. doi:10.1093/molbev/msw209. PMID 27729397.
- ↑ "Biomineralization of limpet teeth: a cryo-TEM study of the organic matrix and the onset of mineral deposition". Journal of Structural Biology 158 (3): 428–444. June 2007. doi:10.1016/j.jsb.2007.01.001. PMID 17306563.
- ↑ "A cover of glass: first report of biomineralized silicon in a ciliate, Maryna umbrellata (Ciliophora: Colpodea)". The Journal of Eukaryotic Microbiology 56 (6): 519–530. 2009. doi:10.1111/j.1550-7408.2009.00431.x. PMID 19883440.
- ↑ 55.0 55.1 "Siliceous structures and silicification in flagellated protists". Protoplasma 181 (1–4): 29–42. 1994. doi:10.1007/BF01666387. Bibcode: 1994Prpls.181...29P.
- ↑ "Role of silicon in diatom metabolism. A silicon requirement for deoxyribonucleic acid synthesis in the diatom Cylindrotheca fusiformis Reimann and Lewin". Experimental Cell Research 58 (2): 334–342. December 1969. doi:10.1016/0014-4827(69)90514-X. PMID 5404077.
- ↑ "Silicon Metabolism in Diatoms: Implications for Growth". Journal of Phycology 36 (5): 821–840. 2000. doi:10.1046/j.1529-8817.2000.00019.x. Bibcode: 2000JPcgy..36..821M.
- ↑ 58.0 58.1 58.2 58.3 58.4 "Competition between Silicifiers and Non-silicifiers in the Past and Present Ocean and Its Evolutionary Impacts". Frontiers in Marine Science 5: 22. 2018. doi:10.3389/fmars.2018.00022. Bibcode: 2018FrMaS...5...22H. 50px Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ Biomineralization: Principles and Concepts in Bioinorganic Materials Chemistry. Oxford University Press. 2001. ISBN 978-0-19-850882-3. https://books.google.com/books?id=YLNl5Jm7XxwC&q=%22Biomineralization.+Principles+and+Concepts+in+Bioinorganic+Materials+Chemistry%22.
- ↑ "The evolution of silicification in diatoms: Inescapable sinking and sinking as escape?". New Phytologist 162 (1): 45–61. 2004. doi:10.1111/j.1469-8137.2004.01022.x. Bibcode: 2004NewPh.162...45R.
- ↑ "Silica Use Through Time: Macroevolutionary Change in the Morphology of the Diatom Fustule". Geomicrobiology Journal 27 (6–7): 596–608. 2010. doi:10.1080/01490451003702941. Bibcode: 2010GmbJ...27..596F.
- ↑ "The Transport and Function of Silicon in Plants". Biological Reviews 58 (2): 179–207. 1983. doi:10.1111/j.1469-185X.1983.tb00385.x.
- ↑ "The amino acid and sugar composition of diatom cell-walls". Marine Biology 19 (4): 323–331. 1973. doi:10.1007/BF00348902. Bibcode: 1973MarBi..19..323H.
- ↑ "Computational model for protein-mediated biomineralization of the diatom frustule". Marine Biology 126 (3): 353–360. 1996. doi:10.1007/BF00354617. Bibcode: 1996MarBi.126..353L.
- ↑ "Architecture and material properties of diatom shells provide effective mechanical protection". Nature 421 (6925): 841–843. February 2003. doi:10.1038/nature01416. PMID 12594512. Bibcode: 2003Natur.421..841H. https://epic.awi.de/id/eprint/5688/1/Ham2002b.pdf.
- ↑ "Microstructure provides insights into evolutionary design and resilience of Coscinodiscus sp. frustule". Proceedings of the National Academy of Sciences of the United States of America 113 (8): 2017–2022. February 2016. doi:10.1073/pnas.1519790113. PMID 26858446. Bibcode: 2016PNAS..113.2017A.
- ↑ "Nanostructural Organization of Naturally Occurring Composites—Part I: Silica-Collagen-Based Biocomposites". Journal of Nanomaterials 2008: 1–8. 2008. doi:10.1155/2008/623838.
- ↑ "Glassin, a histidine-rich protein from the siliceous skeletal system of the marine sponge Euplectella, directs silica polycondensation". Proceedings of the National Academy of Sciences of the United States of America 112 (37): 11449–11454. September 2015. doi:10.1073/pnas.1506968112. PMID 26261346. Bibcode: 2015PNAS..11211449S.
- ↑ "Hierarchical assembly of the siliceous skeletal lattice of the hexactinellid sponge Euplectella aspergillum". Journal of Structural Biology 158 (1): 93–106. April 2007. doi:10.1016/j.jsb.2006.10.027. PMID 17175169.
- ↑ "Silicification of the medial tooth in the blue crab Callinectes sapidus". Journal of Morphology 277 (12): 1648–1660. December 2016. doi:10.1002/jmor.20614. PMID 27650814.
- ↑ "Grazing-induced changes in cell wall silicification in a marine diatom". Protist 158 (1): 21–28. January 2007. doi:10.1016/j.protis.2006.09.002. PMID 17081802.
- ↑ "Size and biomechanic properties of diatom frustules influence food uptake by copepods". Marine Ecology Progress Series 481: 41–51. 2013. doi:10.3354/meps10227. Bibcode: 2013MEPS..481...41F. https://www.int-res.com/abstracts/meps/v481/p41-51/.
- ↑ "The ecology of herbivore-induced silicon defences in grasses". Functional Ecology 30 (8): 1311–1322. 2016. doi:10.1111/1365-2435.12706. Bibcode: 2016FuEco..30.1311H.
- ↑ "UV-screening of grasses by plant silica layer?". Journal of Biosciences 38 (2): 413–416. June 2013. doi:10.1007/s12038-013-9303-1. PMID 23660676.
- ↑ "Diatoms as living photonic crystals". Applied Physics B 78 (3–4): 257–260. 2004. doi:10.1007/s00340-004-1419-4. Bibcode: 2004ApPhB..78..257F.
- ↑ "Optical properties of diatom silica frustule with special reference to blue light". Journal of Applied Physics 103 (7): 074701–074701–5. 2008. doi:10.1063/1.2903342. Bibcode: 2008JAP...103g4701Y.
- ↑ "Wavelength and orientation dependent capture of light by diatom frustule nanostructures". Scientific Reports 5 (1). December 2015. doi:10.1038/srep17403. PMID 26627680. Bibcode: 2015NatSR...517403R.
- ↑ "Fibre-optical features of a glass sponge". Nature 424 (6951): 899–900. August 2003. doi:10.1038/424899a. PMID 12931176. Bibcode: 2003Natur.424..899S.
- ↑ "A dynamic broadband reflector built from microscopic silica spheres in the 'disco' clam Ctenoides ales". Journal of the Royal Society, Interface 11 (98). September 2014. doi:10.1098/rsif.2014.0407. PMID 24966236.
- ↑ "Aluminum-dependent regulation of intracellular silicon in the aquatic invertebrate Lymnaea stagnalis". Proceedings of the National Academy of Sciences of the United States of America 99 (6): 3394–3399. March 2002. doi:10.1073/pnas.062478699. PMID 11891333. Bibcode: 2002PNAS...99.3394D.
- ↑ "Silicon and heavy metal tolerance of higher plants". Phytochemistry 56 (7): 685–692. April 2001. doi:10.1016/S0031-9422(00)00472-6. PMID 11314953. Bibcode: 2001PChem..56..685N.
- ↑ "A proton buffering role for silica in diatoms". Science 297 (5588): 1848–1850. September 2002. doi:10.1126/science.1074958. PMID 12228711. Bibcode: 2002Sci...297.1848M.
- ↑ "The revised classification of eukaryotes". The Journal of Eukaryotic Microbiology 59 (5): 429–493. September 2012. doi:10.1111/j.1550-7408.2012.00644.x. PMID 23020233.
- ↑ "A first report of hydroxylated apatite as structural biomineral in Loasaceae - plants' teeth against herbivores". Scientific Reports 6 (1). May 2016. doi:10.1038/srep26073. PMID 27194462. Bibcode: 2016NatSR...626073E.
- ↑ "Plant cystoliths: a complex functional biocomposite of four distinct silica and amorphous calcium carbonate phases". Chemistry: A European Journal 18 (33): 10262–10270. August 2012. doi:10.1002/chem.201201111. PMID 22696477.
- ↑ "Phosphorus-31 NMR Studies of Cell Wall-Associated Calcium-Phosphates in Ulva lactuca". Plant Physiology 90 (1): 230–236. May 1989. doi:10.1104/pp.90.1.230. PMID 16666741.
- ↑ "Resource Competition between Plankton Algae: An Experimental and Theoretical Approach". Ecology 58 (2): 338–348. 1977. doi:10.2307/1935608. Bibcode: 1977Ecol...58..338T.
- ↑ "The impact of light intensity and daylength on silicate and nitrate competition among marine phytoplankton". Limnology and Oceanography 39 (7): 1680–1688. 1994. doi:10.4319/lo.1994.39.7.1680. Bibcode: 1994LimOc..39.1680S. https://oceanrep.geomar.de/id/eprint/14122/1/Sommer_1994.pdf.
- ↑ "The rise of predators". Geology 39 (6): 607–608. 2011. doi:10.1130/focus062011.1. Bibcode: 2011Geo....39..607P.
- ↑ "Phosphate biomineralization in mid-Neoproterozoic protists". Geology 39 (6): 539–542. 2011. doi:10.1130/G31833.1. Bibcode: 2011Geo....39..539C.
- ↑ "Possible animal-body fossils in pre-Marinoan limestones from South Australia". Nature Geoscience 3 (9): 653–659. 2010. doi:10.1038/ngeo934. Bibcode: 2010NatGe...3..653M.
- ↑ "Proterozoic modular biomineralized metazoan from the Nama Group, Namibia". Science 296 (5577): 2383–2386. June 2002. doi:10.1126/science.1071599. PMID 12089440. Bibcode: 2002Sci...296.2383W.
- ↑ 93.0 93.1 "Eve of biomineralization: Controls on skeletal mineralogy". Geology 36 (12): 923. 2008. doi:10.1130/G25094A.1. Bibcode: 2008Geo....36..923Z. http://wzar.unizar.es/murero/activos/pdfs/2008_Zhuravlev%26Wood_Geology.pdf. Retrieved 28 August 2015.
- ↑ "Seawater chemistry and early carbonate biomineralization". Science 316 (5829): 1302. June 2007. doi:10.1126/science.1137284. PMID 17540895. Bibcode: 2007Sci...316.1302P.
- ↑ "The earliest Cambrian record of animals and ocean geochemical change". Geological Society of America Bulletin 122 (11–12): 1731–1774. 2010. doi:10.1130/B30346.1. Bibcode: 2010GSAB..122.1731M.
- ↑ "Evolutionary origins of animal skeletal biomineralization". Cells Tissues Organs 194 (2–4): 98–102. 2011. doi:10.1159/000324245. PMID 21625061.
- ↑ "Chronology of early Cambrian biomineralization". Geological Magazine 149 (2): 221–251. 2011. doi:10.1017/S0016756811000720. Bibcode: 2012GeoM..149..221K.
- ↑ 98.0 98.1 "A marriage of bone and nacre". Nature 392 (6679): 861–862. April 1998. doi:10.1038/31798. PMID 9582064. Bibcode: 1998Natur.392..861W.
- ↑ 99.0 99.1 "Parallel evolution of nacre building gene sets in molluscs". Molecular Biology and Evolution 27 (3): 591–608. March 2010. doi:10.1093/molbev/msp278. PMID 19915030.
- ↑ "Skeletal matrices, muci, and the origin of invertebrate calcification". Proceedings of the National Academy of Sciences of the United States of America 93 (4): 1554–1559. February 1996. doi:10.1073/pnas.93.4.1554. PMID 11607630. Bibcode: 1996PNAS...93.1554M.
- ↑ "Evolutionary prerequisites for early Phanerozoic calcareous skeletons". Bio Systems 12 (1–2): 27–41. 1980. doi:10.1016/0303-2647(80)90036-2. PMID 6991017. Bibcode: 1980BiSys..12...27L.
- ↑ "Evolutionary prerequisites for early Phanerozoic calcareous skeletons". Bio Systems 12 (1–2): 27–41. 1980. doi:10.1016/0303-2647(80)90036-2. PMID 6991017. Bibcode: 1980BiSys..12...27L.
- ↑ "Differential expression of three galaxin-related genes during settlement and metamorphosis in the scleractinian coral Acropora millepora". BMC Evolutionary Biology 9 (1): 178. July 2009. doi:10.1186/1471-2148-9-178. PMID 19638240. Bibcode: 2009BMCEE...9..178R.
- ↑ "Sponge paleogenomics reveals an ancient role for carbonic anhydrase in skeletogenesis". Science 316 (5833): 1893–1895. June 2007. doi:10.1126/science.1141560. PMID 17540861. Bibcode: 2007Sci...316.1893J.
- ↑ "Brachiopod and mollusc biomineralisation is a conserved process that was lost in the phoronid-bryozoan stem lineage". EvoDevo 13 (1): 17. September 2022. doi:10.1186/s13227-022-00202-8. PMID 36123753.
- ↑ "10 A Grand Unified theory of Biomineralization.". The Biomineralisation of Nano- and Micro-Structures. Weinheim, Germany: Wiley-VCH. 2000. pp. 139–150.
- ↑ "Ultrastructure and development of iron mineralization in the radular teeth of Cryptochiton stelleri (Mollusca)". Journal of Ultrastructure Research 17 (1): 1–13. January 1967. doi:10.1016/S0022-5320(67)80015-7. PMID 6017357.
- ↑ Nanophase and Nanocomposite Materials: Symposium Held December 1–3, 1992, Boston, Massachusetts, U.S.A. (Materials Research Society Symposium Proceedings). Pittsburgh, Pa: Materials Research Society. 1993. ISBN 978-1-55899-181-1.
- ↑ 109.0 109.1 Biomineralization: From Nature to Application. John Wiley & Sons. 30 April 2008. ISBN 978-0-470-98631-8. https://books.google.com/books?id=TiwK2VQhPMkC&q=biomineralization+applications.
- ↑ 110.0 110.1 Biomineralization and Biomaterials: Fundamentals and Applications. Woodhead. 28 September 2015. ISBN 978-1-78242-356-0. https://books.google.com/books?id=UhGpBAAAQBAJ&q=biomineralization+applications.
- ↑ "Microbial sciences: the superficial life of microbes". Nature 441 (7091): 300–302. May 2006. doi:10.1038/441300a. PMID 16710410. Bibcode: 2006Natur.441..300K.
- ↑ "Bacterial cell attachment, the beginning of a biofilm". Journal of Industrial Microbiology & Biotechnology 34 (9): 577–588. September 2007. doi:10.1007/s10295-007-0234-4. PMID 17619090.
- ↑ 113.0 113.1 "Biofilms: the matrix revisited". Trends in Microbiology 13 (1): 20–26. January 2005. doi:10.1016/j.tim.2004.11.006. PMID 15639628.
- ↑ "The Matrix Reloaded: Probing the Extracellular Matrix Synchronizes Bacterial Communities". Journal of Bacteriology 197 (13): 2092–2103. July 2015. doi:10.1128/JB.02516-14. PMID 25825428.
- ↑ "The Peculiar Functions of the Bacterial Extracellular Matrix". Trends in Microbiology 25 (4): 257–266. April 2017. doi:10.1016/j.tim.2016.12.010. PMID 28089324.
- ↑ 116.0 116.1 116.2 116.3 "Architects of nature: growing buildings with bacterial biofilms". Microbial Biotechnology 10 (5): 1157–1163. September 2017. doi:10.1111/1751-7915.12833. PMID 28815998. 50px Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ "Spatio-temporal assembly of functional mineral scaffolds within microbial biofilms". npj Biofilms and Microbiomes 2: 15031. 2016. doi:10.1038/npjbiofilms.2015.31. PMID 28721240. 50px Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ "Self healing concrete: a biological approach". Self Healing Materials: An Alternative Approach to 20 Centuries of Materials Science. Springer. 2007. pp. 195–204. ISBN 978-1-4020-6250-6. https://books.google.com/books?id=v4zDNBtACIsC&q=Self+healing+concrete%3A+a+biological+approach.&pg=PA194.
- ↑ Dosier GK, "Methods for making construction material using enzyme producing bacteria", US patent 8728365, issued 2014, assigned to Biomason Inc.
- ↑ "Osmotic pressure can regulate matrix gene expression in Bacillus subtilis". Molecular Microbiology 86 (2): 426–436. October 2012. doi:10.1111/j.1365-2958.2012.08201.x. PMID 22882172.
- ↑ "Defects in the flagellar motor increase synthesis of poly-γ-glutamate in Bacillus subtilis". Journal of Bacteriology 196 (4): 740–753. February 2014. doi:10.1128/JB.01217-13. PMID 24296669.
- ↑ 122.0 122.1 "The biogeochemistry and bioremediation of uranium and other priority radionuclides". Chemical Geology 363: 164–184. 2014. doi:10.1016/j.chemgeo.2013.10.034. Bibcode: 2014ChGeo.363..164N.
- ↑ Environmental microbe-metal interactions: Bioremediation of radionuclide-containing wastewaters. Washington, DC: ASM Press. 2000. pp. 277–327. ISBN 978-1-55581-195-2.
- ↑ "The New IMA List of Minerals A Work in Progress". The New IMA List of Minerals. IMA – CNMNC (Commission on New Minerals Nomenclature and Classification). November 2020. http://cnmnc.main.jp/IMA_Master_List_%282020-11%29.pdf.
- ↑ "IMA Database of Mineral Properties/ RRUFF Project". Department of Geosciences, University of Arizona. https://rruff.info/ima/.
- ↑ 126.0 126.1 "Minerals formed by organisms". Science 211 (4487): 1126–1131. March 1981. doi:10.1126/science.7008198. PMID 7008198. Bibcode: 1981Sci...211.1126L.
- ↑ 127.0 127.1 127.2 127.3 127.4 "Biominerals". Mineralogical Magazine 69 (5): 621–41. 2005. doi:10.1180/0026461056950275. Bibcode: 2005MinM...69..621S.
- ↑ 128.0 128.1 "The definition of a mineral". The Canadian Mineralogist 33 (3): 689–90. 1995. https://pubs.geoscienceworld.org/canmin/article-abstract/33/3/689/12679/the-definition-of-a-mineral?redirectedFrom=fulltext.
- ↑ "Working Group on Environmental Mineralogy and Geochemistry" (in en). International Mineralogical Association. 3 August 2011. https://www.ima-mineralogy.org/WGEMG_objectives.htm.
- ↑ "Limits of life and the biosphere: Lessons from the detection of microorganisms in the deep sea and deep subsurface of the Earth.". Origins and Evolution of Life: An Astrobiological Perspective. Cambridge: Cambridge University Press. 2010. pp. 469–86. ISBN 978-1-139-49459-5.
- ↑ "Extending the sub-sea-floor biosphere". Science 320 (5879): 1046. May 2008. doi:10.1126/science.1154545. PMID 18497290. Bibcode: 2008Sci...320.1046R. https://archimer.ifremer.fr/doc/00000/4209/.
- ↑ "Microorganisms in the atmosphere over Antarctica". FEMS Microbiology Ecology 69 (2): 143–157. August 2009. doi:10.1111/j.1574-6941.2009.00706.x. PMID 19527292. Bibcode: 2009FEMME..69..143P.
- ↑ "Geomicrobiology: how molecular-scale interactions underpin biogeochemical systems". Science 296 (5570): 1071–1077. May 2002. doi:10.1126/science.1010716. PMID 12004119. Bibcode: 2002Sci...296.1071N.
- ↑ "Geoscience. Microbial geoengineers". Science 299 (5609): 1027–1029. February 2003. doi:10.1126/science.1072076. PMID 12586932.
- ↑ "Bacterial biomineralization: new insights from Myxococcus-induced mineral precipitation". Geological Society, London, Special Publications 336 (1): 31–50. 2010. doi:10.1144/SP336.3. Bibcode: 2010GSLSP.336...31G.
- ↑ "Biomineralization. Cell Biology and Mineral Deposition. by Kenneth Simkiss; Karl M. Wilbur On Biomineralization. by Heinz A. Lowenstam; Stephen Weiner". Science 247 (4946): 1129–30. 1990. doi:10.1126/science.247.4946.1129. PMID 17800080. Bibcode: 1990Sci...247.1129S.
- ↑ "Official IMA list of mineral names". uws.edu.au. March 2009. http://pubsites.uws.edu.au/ima-cnmnc/IMA%20mineral%20list%20update%20BB%20Upload%208%20April%202011.pdf.
- ↑ Earth Materials. Wiley-Blackwell. 2010. ISBN 978-1-4443-3460-9.
- ↑ "Icosahedrite, Al63Cu24Fe13, the first natural quasicrystal". American Mineralogist 96 (5–6): 928–31. 2011. doi:10.2138/am.2011.3758. Bibcode: 2011AmMin..96..928B.
- ↑ "Approved as new mineral". Commission on New Minerals and Mineral Names. http://pubsites.uws.edu.au/ima-cnmnc/newminerals2010.pdf.
- ↑ "Biogenic Minerals". Science Frontiers 66. Nov–Dec 1989. https://www.science-frontiers.com/sf066/sf066g13.htm.
- ↑ He, Honghua; Veneklaas, Erik J.; Kuo, John; Lambers, Hans (2014-03-01). "Physiological and ecological significance of biomineralization in plants". Trends in Plant Science 19 (3): 166–174. doi:10.1016/j.tplants.2013.11.002. ISSN 1360-1385. PMID 24291440. Bibcode: 2014TPS....19..166H. https://www.sciencedirect.com/science/article/abs/pii/S1360138513002537.
- ↑ "Final report of the MEPAG Astrobiology Field Laboratory Science Steering Group (AFL-SSG)". The Astrobiology Field Laboratory. U.S.A.: Mars Exploration Program Analysis Group (MEPAG) - NASA. 26 September 2006. p. 72. http://mepag.jpl.nasa.gov/reports/AFL_SSG_WHITE_PAPER_v3.doc. Retrieved 22 July 2009.
- ↑ 144.0 144.1 "Exploring martian habitability. Habitability, taphonomy, and the search for organic carbon on Mars. Introduction". Science 343 (6169): 386–387. January 2014. doi:10.1126/science.1249944. PMID 24458635. Bibcode: 2014Sci...343..386G.
- ↑ 145.0 145.1 Various (24 January 2014). "Special Issue - Table of Contents - Exploring Martian Habitability". Science 343: 345–452. https://www.science.org/toc/science/343/6169. Retrieved 24 January 2014.
- ↑ Various (24 January 2014). "Special Collection - Curiosity - Exploring Martian Habitability". Science. https://www.science.org/action/doSearch?AllField=Curiosity+Mars. Retrieved 24 January 2014.
- ↑ "A habitable fluvio-lacustrine environment at Yellowknife Bay, Gale crater, Mars". Science 343 (6169). January 2014. doi:10.1126/science.1242777. PMID 24324272. Bibcode: 2014Sci...343A.386G.
Further reading
- "Control And Design Principles In Biological Mineralization" (abstract). Angewandte Chemie International Edition in English 31 (2): 153–169. 1992. doi:10.1002/anie.199201531. https://onlinelibrary.wiley.com/doi/10.1002/anie.199201531.
- "Biomineralization: an overview". Connective Tissue Research 44 (Supplement 1): 5–9. 2003. doi:10.1080/713713622. PMID 12952166.
- "Biomineralization and diagenesis in the Scleractinia: part I, biomineralization". Bull. Tohoku Univ. Museum 1: 144–151. 2001.
- "Structures, organo mineral compositions and diagenetic changes in biominerals". Current Opinion in Colloid & Interface Science 7 (1–2): 133–138. 2002. doi:10.1016/S1359-0294(02)00013-4.
- "Biomineralization". Encyclopedia of Inorganic Chemistry (Wiley & Sons) 1: 391–404. 2005. ISBN 978-0-521-87473-1.
- "Serpulids living deep: calcareous tubeworms beyond the abyss". Deep-Sea Research Part I 90: 91–104. 2014. doi:10.1016/j.dsr.2014.04.006. Bibcode: 2014DSRI...90...91K. https://www.academia.edu/8904126. Retrieved 9 January 2014.
- "Minerals formed by organisms". Science 211 (4487): 1126–1131. March 1981. doi:10.1126/science.7008198. PMID 7008198. Bibcode: 1981Sci...211.1126L.
- "The Little Workers of the Mining Industry". Science Creative Quarterly (2). 2006. https://www.scq.ubc.ca/the-little-workers-of-the-mining-industry/. Retrieved 3 November 2006.
- "Micro-environmental controls on biomineralization: superficial processes of apatite and calcite precipitation in Quaternary soils, Roussillon, France". Sedimentology 46 (3): 463–476. 1999. doi:10.1046/j.1365-3091.1999.00224.x. Bibcode: 1999Sedim..46..463S.
- "The Formation of Iron Biominerals". Metals, Microbes, and Minerals - The Biogeochemical Side of Life. Berlin: De Gruyter. 2021. pp. 159–184. doi:10.1515/9783110589771-006. ISBN 978-3-11-058977-1.
- "Occurrence, formation and function of organic sheets in the mineral tube structures of Serpulidae (polychaeta, Annelida)". PLOS ONE 8 (10). 2013. doi:10.1371/journal.pone.0075330. PMID 24116035. Bibcode: 2013PLoSO...875330V.
- "Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida)". Zoological Journal of the Linnean Society 154 (4): 633–650. 2008. doi:10.1111/j.1096-3642.2008.00421.x. https://www.researchgate.net/publication/222089798. Retrieved 9 January 2014.
- "Design strategies in mineralized biological materials". Journal of Materials Chemistry 7 (5): 689–702. 1997. doi:10.1039/a604512j.
External links
- 'Data and literature on modern and fossil Biominerals'
- The Little Workers of the Mining Industry (an overview of the bacteria involved in biomineralization), Science Creative Quarterly
- Biomineralization web-book: bio-mineral.org
- Minerals and the Origins of Life (Robert Hazen, NASA) (video, 60m, April 2014).
- Special German Research Project About the Principles of Biomineralization
 |  |
