Biology:Neutrophil
| Neutrophil | |
|---|---|
.png) 3D rendering of a neutrophil | |
 Neutrophils with segmented nuclei surrounded by erythrocytes and platelets. Intra-cellular granules are visible in the cytoplasm (Giemsa stained). | |
| Details | |
| System | Immune system |
| Function | Phagocytosis |
| Anatomical terms of microanatomy | |
Neutrophils (also known as neutrocytes, heterophils or polymorphonuclear leukocytes) are a type of white blood cell. More specifically, they form the most abundant type of granulocytes and make up 40% to 70% of all white blood cells in humans.[1] They form an essential part of the innate immune system, with their functions varying in different animals.[2]
They are formed from stem cells in the bone marrow and differentiated into subpopulations of neutrophil-killers and neutrophil-cagers. They are short-lived (between 5 and 135 hours, see § Life span) and highly mobile, as they can enter parts of tissue where other cells/molecules cannot. Neutrophils may be subdivided into segmented neutrophils and banded neutrophils (or bands). They form part of the polymorphonuclear cells family (PMNs) together with basophils and eosinophils.[3][4][5]
The name neutrophil derives from staining characteristics on hematoxylin and eosin (H&E) histological or cytological preparations. Whereas basophilic white blood cells stain dark blue and eosinophilic white blood cells stain bright red, neutrophils stain a neutral pink. Normally, neutrophils contain a nucleus divided into 2–5 lobes.[6]
Neutrophils are a type of phagocyte and are normally found in the bloodstream. During the beginning (acute) phase of inflammation, particularly as a result of bacterial infection, environmental exposure,[7] and some cancers,[8][9] neutrophils are one of the first responders of inflammatory cells to migrate toward the site of inflammation. They migrate through the blood vessels and then through interstitial space, following chemical signals such as interleukin-8 (IL-8), C5a, fMLP, leukotriene B4, and hydrogen peroxide (H2O2)[10] in a process called chemotaxis. They are the predominant cells in pus, accounting for its whitish/yellowish appearance.[11]
Neutrophils are recruited to the site of injury within minutes following trauma and are the hallmark of acute inflammation;[12] however, due to some pathogens being indigestible, they might not be able to resolve certain infections without the assistance of other types of immune cells.
Structure


When adhered to a surface, neutrophil granulocytes have an average diameter of 12–15 micrometers (µm) in peripheral blood smears. In suspension, human neutrophils have an average diameter of 8.85 µm.[13]
With the eosinophil and the basophil, they form the class of polymorphonuclear cells, named for the nucleus' multilobulated shape (as compared to lymphocytes and monocytes, the other types of white cells). The nucleus has a characteristic lobed appearance, the separate lobes connected by chromatin. The nucleolus disappears as the neutrophil matures, which is something that happens in only a few other types of nucleated cells.[14]:168 Up to 17% of female human neutrophil nuclei have a drumstick-shaped appendage which contains the inactivated X chromosome.[15] In the cytoplasm, the Golgi apparatus is small, mitochondria and ribosomes are sparse, and the rough endoplasmic reticulum is absent.[14]:170 The cytoplasm also contains about 200 granules, of which a third are azurophilic.[14]:170
Neutrophils will show increasing segmentation (many segments of the nucleus) as they mature. A normal neutrophil should have 3–5 segments. Hypersegmentation is not normal but occurs in some disorders, most notably vitamin B12 deficiency. This is noted in a manual review of the blood smear and is positive when most or all of the neutrophils have 5 or more segments.

Neutrophils are the most abundant white blood cells in humans (approximately 1011 are produced daily); they account for approximately 50–70% of all white blood cells (leukocytes). The stated normal range for human blood counts varies between laboratories, but a neutrophil count of 2.5–7.5 × 109/L is a standard normal range. People of African and Middle Eastern descent may have lower counts, which are still normal.[16] A report may divide neutrophils into segmented neutrophils and bands.
When circulating in the bloodstream and inactivated, neutrophils are spherical. Once activated, they change shape and become more amorphous or amoeba-like and can extend pseudopods as they hunt for antigens.[17]
In 1973, Sanchez et al. found that the capacity of neutrophils to engulf bacteria is reduced when simple sugars like glucose, fructose as well as sucrose, honey and orange juice were ingested, while the ingestion of starches had no effect. Fasting, on the other hand, strengthened the neutrophils' phagocytic capacity to engulf bacteria. It was concluded that the function, and not the number, of phagocytes in engulfing bacteria was altered by the ingestion of sugars.[18] In 2007 researchers at the Whitehead Institute of Biomedical Research found that given a selection of sugars on microbial surfaces, the neutrophils reacted to some types of sugars preferentially. The neutrophils preferentially engulfed and killed beta-1,6-glucan targets compared to beta-1,3-glucan targets.[19][20]
Development
Life span

The average lifespan of inactivated human neutrophils in the circulation has been reported by different approaches to be between 5 and 135 hours(5 days and 15 hours).[21][22]
Upon activation, they marginate (position themselves adjacent to the blood vessel endothelium) and undergo selectin-dependent capture followed by integrin-dependent adhesion in most cases, after which they migrate into tissues, where they survive for 1–2 days.[23] Neutrophils have also been demonstrated to be released into the blood from a splenic reserve following myocardial infarction.[24]
Neutrophils are much more numerous than the longer-lived monocyte/macrophage phagocytes. A pathogen (disease-causing microorganism or virus) is likely to first encounter a neutrophil. Some experts hypothesize that the short lifetime of neutrophils is an evolutionary adaptation. The short lifetime of neutrophils minimizes propagation of those pathogens that parasitize phagocytes (e.g. Leishmania[25]) because the more time such parasites spend outside a host cell, the more likely they will be destroyed by some component of the body's defenses. Also, because neutrophil antimicrobial products can also damage host tissues, their short life limits damage to the host during inflammation.[23]
Neutrophils will be removed after phagocytosis of pathogens by macrophages. PECAM-1 and phosphatidylserine on the cell surface are involved in this process.
Function
Chemotaxis
Neutrophils undergo a process called chemotaxis via amoeboid movement, which allows them to migrate toward sites of infection or inflammation. Cell surface receptors allow neutrophils to detect chemical gradients of molecules such as interleukin-8 (IL-8), interferon gamma (IFN-γ), C3a, C5a, and leukotriene B4, which these cells use to direct the path of their migration.
Neutrophils have a variety of specific receptors, including ones for complement, cytokines like interleukins and IFN-γ, chemokines, lectins, and other proteins. They also express receptors to detect and adhere to endothelium and Fc receptors for opsonin.[26]
In leukocytes responding to a chemoattractant, the cellular polarity is regulated by activities of small Rho guanosine triphosphatases (Rho GTPases) and the phosphoinositide 3-kinases (PI3Ks). In neutrophils, lipid products of PI3Ks regulate activation of Rac1, hematopoietic Rac2, and RhoG GTPases of the Rho family and are required for cell motility. Rac-GTPases regulate cytoskeletal dynamics and facilitate neutrophils adhesion, migration, and spreading.[27] They accumulate asymmetrically to the plasma membrane at the leading edge of polarized cells. Spatially regulating Rho GTPases and organizing the leading edge of the cell, PI3Ks and their lipid products could play pivotal roles in establishing leukocyte polarity, as compass molecules that tell the cell where to crawl.
It has been shown in mice that in certain conditions neutrophils have a specific type of migration behaviour referred to as neutrophil swarming during which they migrate in a highly coordinated manner and accumulate and cluster to sites of inflammation.[28]
Anti-microbial function
Being highly motile, neutrophils quickly congregate at a focus of infection, attracted by cytokines expressed by activated endothelium, mast cells, and macrophages. Neutrophils express[29] and release cytokines, which in turn amplify inflammatory reactions by several other cell types.
In addition to recruiting and activating other cells of the immune system, neutrophils play a key role in the front-line defense against invading pathogens. Neutrophils have three methods for directly attacking micro-organisms: phagocytosis (ingestion), degranulation (release of soluble anti-microbials), and generation of neutrophil extracellular traps (NETs).[30]
Phagocytosis

Neutrophils are phagocytes, capable of ingesting microorganisms or particles. For targets to be recognized, they must be coated in opsonins – a process known as antibody opsonization.[17] They can internalize and kill many microbes, each phagocytic event resulting in the formation of a phagosome into which reactive oxygen species and hydrolytic enzymes are secreted. The consumption of oxygen during the generation of reactive oxygen species has been termed the "respiratory burst", although unrelated to respiration or energy production.
The respiratory burst involves the activation of the enzyme NADPH oxidase, which produces large quantities of superoxide, a reactive oxygen species. Superoxide decays spontaneously or is broken down via enzymes known as superoxide dismutases (Cu/ZnSOD and MnSOD), to hydrogen peroxide, which is then converted to hypochlorous acid (HClO), by the green heme enzyme myeloperoxidase. It is thought that the bactericidal properties of HClO are enough to kill bacteria phagocytosed by the neutrophil, but this may instead be a step necessary for the activation of proteases.[31]
Though neutrophils can kill many microbes, the interaction of neutrophils with microbes and molecules produced by microbes often alters neutrophil turnover. The ability of microbes to alter the fate of neutrophils is highly varied, can be microbe-specific, and ranges from prolonging the neutrophil lifespan to causing rapid neutrophil lysis after phagocytosis. Chlamydia pneumoniae and Neisseria gonorrhoeae have been reported to delay neutrophil apoptosis.[32][33][34] Thus, some bacteria – and those that are predominantly intracellular pathogens – can extend the neutrophil lifespan by disrupting the normal process of spontaneous apoptosis and/or PICD (phagocytosis-induced cell death). On the other end of the spectrum, some pathogens such as Streptococcus pyogenes are capable of altering neutrophil fate after phagocytosis by promoting rapid cell lysis and/or accelerating apoptosis to the point of secondary necrosis.[35][36]
Degranulation
Neutrophils also release an assortment of proteins in three types of granules by a process called degranulation. The contents of these granules have antimicrobial properties, and help combat infection. Glitter cells are polymorphonuclear leukocyte neutrophils with granules.[37]
| Granule type | Protein |
| Azurophilic granules (or "primary granules") | Myeloperoxidase, bactericidal/permeability-increasing protein (BPI), defensins, and the serine proteases neutrophil elastase, Proteinase 3 and cathepsin G |
| Specific granules (or "secondary granules") | Alkaline phosphatase, lysozyme, NADPH oxidase, collagenase, lactoferrin, histaminase,[38] and cathelicidin |
| Tertiary granules | Cathepsin, gelatinase, and collagenase |
Neutrophil extracellular traps
In 2004, Brinkmann and colleagues described a striking observation that activation of neutrophils causes the release of web-like structures of DNA; this represents a third mechanism for killing bacteria.[39] These neutrophil extracellular traps (NETs) comprise a web of fibers composed of chromatin and serine proteases [40] that trap and kill extracellular microbes. It is suggested that NETs provide a high local concentration of antimicrobial components and bind, disarm, and kill microbes independent of phagocytic uptake. In addition to their possible antimicrobial properties, NETs may serve as a physical barrier that prevents further spread of pathogens. Trapping of bacteria may be a particularly important role for NETs in sepsis, where NETs are formed within blood vessels.[41] Finally, NET formation has been demonstrated to augment macrophage bactericidal activity during infection.[42][43] Recently, NETs have been shown to play a role in inflammatory diseases, as NETs could be detected in preeclampsia, a pregnancy-related inflammatory disorder in which neutrophils are known to be activated.[44] Neutrophil NET formation may also impact cardiovascular disease, as NETs may influence thrombus formation in coronary arteries.[45][46] NETs are now known to exhibit pro-thrombotic effects both in vitro[47] and in vivo.[48][49] More recently, in 2020 NETs were implicated in the formation of blood clots in cases of severe COVID-19.[50]
Tumor Associated Neutrophils
TANs can exhibit an elevated extracellular acidification rate when there is an increase in glycolysis levels.[51] When there is a metabolic shift in TANS this can lead to tumor progression in certain areas of the body, such as the lungs. In 2021, Ancey et al. identified how TANs support the growth and progression of tumors unlike normal neutrophils which would inhibit tumor progression through the phagocytosis of tumor cells. Utilizing a mouse model, they identified that both Glut1 and glucose metabolism increased in TANs found within a mouse who possessed lung adenocarcinoma.[51]
Clinical significance

Low neutrophil counts are termed neutropenia. This can be congenital (developed at or before birth) or it can develop later, as in the case of aplastic anemia or some kinds of leukemia. It can also be a side-effect of medication, most prominently chemotherapy. Neutropenia makes an individual highly susceptible to infections. It can also be the result of colonization by intracellular neutrophilic parasites.
In alpha 1-antitrypsin deficiency, the important neutrophil elastase is not adequately inhibited by alpha 1-antitrypsin, leading to excessive tissue damage in the presence of inflammation – the most prominent one being emphysema. Negative effects of elastase have also been shown in cases when the neutrophils are excessively activated (in otherwise healthy individuals) and release the enzyme in extracellular space. Unregulated activity of neutrophil elastase can lead to disruption of pulmonary barrier showing symptoms corresponding with acute lung injury.[52] The enzyme also influences activity of macrophages by cleaving their toll-like receptors (TLRs) and downregulating cytokine expression by inhibiting nuclear translocation of NF-κB.[53]
In Familial Mediterranean fever (FMF), a mutation in the pyrin (or marenostrin) gene, which is expressed mainly in neutrophil granulocytes, leads to a constitutively active acute-phase response and causes attacks of fever, arthralgia, peritonitis, and – eventually – amyloidosis.[54]
Hyperglycemia can lead to neutrophil dysfunction. Dysfunction in the neutrophil biochemical pathway myeloperoxidase as well as reduced degranulation are associated with hyperglycemia.[55]
The Absolute neutrophil count (ANC) is also used in diagnosis and prognosis. ANC is the gold standard for determining severity of neutropenia, and thus neutropenic fever. Any ANC < 1500 cells / mm3 is considered neutropenia, but <500 cells / mm3 is considered severe.[56] There is also new research tying ANC to myocardial infarction as an aid in early diagnosis.[57][58] Neutrophils promote ventricular tachycardia in acute myocardial infarction.[59]
In autopsy, the presence of neutrophils in the heart or brain is one of the first signs of infarction, and is useful in the timing and diagnosis of myocardial infarction and stroke.
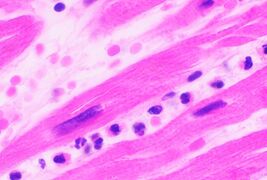
Neutrophils are seen in a myocardial infarction at approximately 12–24 hours,[60] as seen in this micrograph.


Neutrophil antigens
There are five (HNA 1–5) sets of neutrophil antigens recognized.[62] The three HNA-1 antigens (a-c) are located on the low affinity Fc-γ receptor IIIb (FCGR3B :CD16b) The single known HNA-2a antigen is located on CD177. The HNA-3 antigen system has two antigens (3a and 3b) which are located on the seventh exon of the CLT2 gene (SLC44A2). The HNA-4 and HNA-5 antigen systems each have two known antigens (a and b) and are located in the β2 integrin. HNA-4 is located on the αM chain (CD11b) and HNA-5 is located on the αL integrin unit (CD11a).
Subpopulations

Two functionally unequal subpopulations of neutrophils were identified on the basis of different levels of their reactive oxygen metabolite generation, membrane permeability, activity of enzyme system, and ability to be inactivated. The cells of one subpopulation with high membrane permeability (neutrophil-killers) intensively generate reactive oxygen metabolites and are inactivated in consequence of interaction with the substrate, whereas cells of another subpopulation (neutrophil-cagers) produce reactive oxygen species less intensively, don't adhere to substrate and preserve their activity.[63][64][65][66][67] Additional studies have shown that lung tumors can be infiltrated by various populations of neutrophils.[68]
Video

A rapidly moving neutrophil can be seen taking up several conidia over an imaging time of 2 hours with one frame every 30 seconds.
A neutrophil can be seen here selectively taking up several Candida yeasts (fluorescently labeled in green) despite several contacts with Aspergillus fumigatus conidia (unlabeled, white/clear) in a 3-D collagen matrix. Imaging time was 2 hours with one frame every 30 seconds.
Neutrophils display highly directional amoeboid motility in infected footpad and phalanges. Intravital imaging was performed in the footpad path of LysM-eGFP mice 20 minutes after infection with Listeria monocytogenes.[69]
Additional images
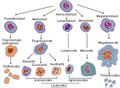
Blood cell lineage

More complete lineages

_diagram_en.svg)
See also
- List of distinct cell types in the adult human body
References
- ↑ Elsevier's Integrated Review Immunology and Microbiology (Second ed.). 2012. doi:10.1016/B978-0-323-07447-6.00002-8.
- ↑ "Candida albicans escapes from mouse neutrophils". Journal of Leukocyte Biology 94 (2): 223–236. August 2013. doi:10.1189/jlb.0213063. PMID 23650619.
- ↑ "Neutrophils: molecules, functions and pathophysiological aspects". Laboratory Investigation; A Journal of Technical Methods and Pathology 80 (5): 617–653. May 2000. doi:10.1038/labinvest.3780067. PMID 10830774.
- ↑ The Neutrophil: Function and Clinical Disorders. Elsevier/North-Holland Amsterdam. 1978. ISBN 978-0-444-80020-6.
- ↑ "Neutrophils and immunity: challenges and opportunities". Nature Reviews. Immunology 6 (3): 173–182. March 2006. doi:10.1038/nri1785. PMID 16498448.
- ↑ Welsh, Charles J. (2021). Hole's Essentials of Human Anatomy and Physiology (14 ed.). New York, USA: McGraw Hill. p. 336. ISBN 978-1-260-57521-7. https://www.mheducation.com/highered/product/hole-s-essentials-human-anatomy-physiology-welsh/M9781260251340.html. Retrieved 28 February 2023.
- ↑ "Subclinical responses in healthy cyclists briefly exposed to traffic-related air pollution: an intervention study". Environmental Health 9 (64): 64. October 2010. doi:10.1186/1476-069X-9-64. PMID 20973949.
- ↑ "The interleukin-8 pathway in cancer". Clinical Cancer Research 14 (21): 6735–6741. November 2008. doi:10.1158/1078-0432.CCR-07-4843. PMID 18980965.
- ↑ "The potential role of neutrophils in promoting the metastatic phenotype of tumors releasing interleukin-8". Clinical Cancer Research 10 (15): 4895–4900. August 2004. doi:10.1158/1078-0432.CCR-03-0760. PMID 15297389.
- ↑ "Lyn is a redox sensor that mediates leukocyte wound attraction in vivo". Nature 480 (7375): 109–112. November 2011. doi:10.1038/nature10632. PMID 22101434. Bibcode: 2011Natur.480..109Y.
- ↑ "The natural history of infection". Medical Microbiology. Elsevier. 2012. pp. 168–173. doi:10.1016/b978-0-7020-4089-4.00029-9. ISBN 978-0-7020-4089-4.
- ↑ Pathways of the Pulp (8th ed.). St. Louis: Mosby. 2002. p. 465.
- ↑ "Trace element landscape of resting and activated human neutrophils on the sub-micrometer level". Metallomics 7 (6): 996–1010. June 2015. doi:10.1039/c4mt00346b. PMID 25832493.
- ↑ 14.0 14.1 14.2 "Neutrophils". Atlas of Blood Cells: Function and Pathology. 1 (2nd ed.). Philadelphia: Lea & Ferbiger. 1988. ISBN 978-0-8121-1094-4.
- ↑ "Nonrandom location and orientation of the inactive X chromosome in human neutrophil nuclei". Chromosoma 110 (4): 267–274. August 2001. doi:10.1007/s004120100145. PMID 11534818.
- ↑ "Reduced neutrophil count in people of African descent is due to a regulatory variant in the Duffy antigen receptor for chemokines gene". PLOS Genetics 5 (1): e1000360. January 2009. doi:10.1371/journal.pgen.1000360. PMID 19180233.
- ↑ 17.0 17.1 Biochemistry and physiology of the neutrophil. Cambridge University Press. 1994. pp. 6. ISBN 978-0-521-41698-6.
- ↑ "Role of sugars in human neutrophilic phagocytosis". The American Journal of Clinical Nutrition 26 (11): 1180–1184. November 1973. doi:10.1093/ajcn/26.11.1180. PMID 4748178. "These data suggest that the function and not the number of phagocytes was altered by ingestion of sugars. This implicates glucose and other simple carbohydrates in the control of phagocytosis and shows that the effects last for at least 5 hr. On the other hand, a fast of 36 or 60 hr significantly increased (P < 0.001) the phagocytic index".
- ↑ "Phagocytosis by human neutrophils is stimulated by a unique fungal cell wall component". Cell Host & Microbe 2 (1): 55–67. July 2007. doi:10.1016/j.chom.2007.06.002. PMID 18005717.
- ↑ "White blood cells are picky about sugar". Whitehead Institute. 2007. http://wi.mit.edu/news/archive/2007/white-blood-cells-are-picky-about-sugar.
- ↑ "What's your age again? Determination of human neutrophil half-lives revisited". Journal of Leukocyte Biology 94 (4): 595–601. October 2013. doi:10.1189/jlb.1112571. PMID 23625199.
- ↑ "In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days". Blood 116 (4): 625–627. July 2010. doi:10.1182/blood-2010-01-259028. PMID 20410504.
- ↑ 23.0 23.1 Wheater's basic histopathology: a colour atlas and text. Edinburgh: Churchill Livingstone. 2002. ISBN 978-0-443-07001-3.
- ↑ Akbar, Naveed; Braithwaite, Adam T; Corr, Emma M; Koelwyn, Graeme J; van Solingen, Coen; Cochain, Clément; Saliba, Antoine-Emmanuel; Corbin, Alastair et al. (4 February 2022). "Rapid neutrophil mobilization by VCAM-1+ endothelial cell-derived extracellular vesicles". Cardiovascular Research 119: 236–251. doi:10.1093/cvr/cvac012. PMID 35134856.
- ↑ Ritter, Uwe; Frischknecht, Freddy; van Zandbergen, Ger (2009). "Are neutrophils important host cells for Leishmania parasites?" (in en). Trends in Parasitology 25 (11): 505–510. doi:10.1016/j.pt.2009.08.003. https://linkinghub.elsevier.com/retrieve/pii/S1471492209001792.
- ↑ Fundamentals of Inflammation. Cambridge University Press. 2010. pp. 53–54. ISBN 978-0-521-88729-8. https://books.google.com/books?id=cJq1RMPKEYkC&q=receptors+neutrophils&pg=PA54.
- ↑ "Rac-GTPases and Rac-GEFs in neutrophil adhesion, migration and recruitment". European Journal of Clinical Investigation 48 (Suppl 2): e12939. November 2018. doi:10.1111/eci.12939. PMID 29682742.
- ↑ "Neutrophil swarms require LTB4 and integrins at sites of cell death in vivo". Nature 498 (7454): 371–375. June 2013. doi:10.1038/nature12175. PMID 23708969. Bibcode: 2013Natur.498..371L.
- ↑ "Cytokine generation, promoter activation, and oxidant-independent NF-kappaB activation in a transfectable human neutrophilic cellular model". BMC Immunology 9: 14. April 2008. doi:10.1186/1471-2172-9-14. PMID 18405381.
- ↑ "Intravascular immunity: the host-pathogen encounter in blood vessels". Nature Reviews. Immunology 9 (5): 364–375. May 2009. doi:10.1038/nri2532. PMID 19390567.
- ↑ "How neutrophils kill microbes". Annual Review of Immunology 23 (5): 197–223. 2005. doi:10.1146/annurev.immunol.23.021704.115653. PMID 15771570.
- ↑ "Neisseria gonorrhoeae delays the onset of apoptosis in polymorphonuclear leukocytes". Cellular Microbiology 8 (11): 1780–1790. November 2006. doi:10.1111/j.1462-5822.2006.00748.x. PMID 16803582.
- ↑ "Neisseria gonorrhoeae-mediated inhibition of apoptotic signalling in polymorphonuclear leukocytes". Infection and Immunity 79 (11): 4447–4458. November 2011. doi:10.1128/IAI.01267-10. PMID 21844239.
- ↑ "Chlamydia pneumoniae multiply in neutrophil granulocytes and delay their spontaneous apoptosis". Journal of Immunology 172 (3): 1768–1776. February 2004. doi:10.4049/jimmunol.172.3.1768. PMID 14734760.
- ↑ "Bacterial pathogens modulate an apoptosis differentiation program in human neutrophils". Proceedings of the National Academy of Sciences of the United States of America 100 (19): 10948–10953. September 2003. doi:10.1073/pnas.1833375100. PMID 12960399.
- ↑ "Influence of Microbes on Neutrophil Life and Death". Frontiers in Cellular and Infection Microbiology 7 (159): 159. 2017. doi:10.3389/fcimb.2017.00159. PMID 28507953.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- ↑ Berman, L. B.; Schreiner, G. E.; Feys, J. O. (1956-11-22). "Observations on the Glitter-Cell Phenomenon" (in en). New England Journal of Medicine 255 (21): 989–991. doi:10.1056/NEJM195611222552104. ISSN 0028-4793. PMID 13378597. http://www.nejm.org/doi/abs/10.1056/NEJM195611222552104.
- ↑ "Localization of histaminase to the specific granule of the human neutrophil". Immunology 52 (4): 649–658. August 1984. PMID 6430792.
- ↑ "Neutrophil extracellular traps kill bacteria". Science 303 (5663): 1532–1535. March 2004. doi:10.1126/science.1092385. PMID 15001782. Bibcode: 2004Sci...303.1532B.
- ↑ "Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans". PLOS Pathogens 5 (10): e1000639. October 2009. doi:10.1371/journal.ppat.1000639. PMID 19876394.
- ↑ "Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood". Nature Medicine 13 (4): 463–469. April 2007. doi:10.1038/nm1565. PMID 17384648.
- ↑ Monteith, Andrew J.; Miller, Jeanette M.; Maxwell, C. Noel; Chazin, Walter J.; Skaar, Eric P. (2021-09-10). "Neutrophil extracellular traps enhance macrophage killing of bacterial pathogens" (in en). Science Advances 7 (37): eabj2101. doi:10.1126/sciadv.abj2101. ISSN 2375-2548. PMID 34516771. Bibcode: 2021SciA....7.2101M.
- ↑ Monteith, Andrew J.; Miller, Jeanette M.; Beavers, William N.; Maloney, K. Nichole; Seifert, Erin L.; Hajnoczky, Gyorgy; Skaar, Eric P. (2021-12-06). "Mitochondrial calcium uniporter affects neutrophil bactericidal activity during Staphylococcus aureus infection" (in en). Infection and Immunity 90 (2): IAI.00551–21. doi:10.1128/IAI.00551-21. ISSN 0019-9567. PMID 34871043.
- ↑ "Neutrophil NETs: a novel contributor to preeclampsia-associated placental hypoxia?". Seminars in Immunopathology 29 (2): 163–167. June 2007. doi:10.1007/s00281-007-0073-4. PMID 17621701. http://doc.rero.ch/record/317255/files/281_2007_Article_73.pdf.
- ↑ "Neutrophil contributions to ischaemic heart disease". European Heart Journal 38 (7): 465–472. February 2017. doi:10.1093/eurheartj/ehx017. PMID 28363210.
- ↑ "Coronary neutrophil extracellular trap burden and deoxyribonuclease activity in ST-elevation acute coronary syndrome are predictors of ST-segment resolution and infarct size". Circulation Research 116 (7): 1182–1192. March 2015. doi:10.1161/CIRCRESAHA.116.304944. PMID 25547404.
- ↑ "Extracellular DNA traps promote thrombosis". Proceedings of the National Academy of Sciences of the United States of America 107 (36): 15880–15885. September 2010. doi:10.1073/pnas.1005743107. PMID 20798043. Bibcode: 2010PNAS..10715880F.
- ↑ "Neutrophil extracellular traps promote deep vein thrombosis in mice". Journal of Thrombosis and Haemostasis 10 (1): 136–144. January 2012. doi:10.1111/j.1538-7836.2011.04544.x. PMID 22044575.
- ↑ "From neutrophil extracellular traps release to thrombosis: an overshooting host-defense mechanism?". Journal of Thrombosis and Haemostasis 9 (9): 1791–1794. September 2011. doi:10.1111/j.1538-7836.2011.04425.x. PMID 21718435.
- ↑ "Neutrophil extracellular traps in COVID-19". Journal of Clinical Investigation 5 (11): e138999. 2020. doi:10.1172/jci.insight.138999. PMID 32329756. PMC 7308057. https://insight.jci.org/articles/view/138999. Retrieved 19 February 2022.
- ↑ 51.0 51.1 Ancey, Pierre-Benoit; Contat, Caroline; Boivin, Gael; Sabatino, Silvia; Pascual, Justine; Zangger, Nadine; Perentes, Jean Yannis; Peters, Solange et al. (March 22, 2021). "Glut1 expression in tumor-associated neutrophils promotes lung cancer growth and resistance to radiotherapy". Cancer Research 81 (9): 2345–2357. doi:10.1158/0008-5472.CAN-20-2870. PMID 33753374.
- ↑ "The role of neutrophil elastase in acute lung injury". European Journal of Pharmacology 451 (1): 1–10. September 2002. doi:10.1016/S0014-2999(02)02182-9. PMID 12223222.
- ↑ "Neutrophil Elastase Subverts the Immune Response by Cleaving Toll-Like Receptors and Cytokines in Pneumococcal Pneumonia". Frontiers in Immunology 9: 732. 2018. doi:10.3389/fimmu.2018.00732. PMID 29922273.
- ↑ "Familial mediterranean fever: revisiting an ancient disease". European Journal of Pediatrics 162 (7–8): 449–454. July 2003. doi:10.1007/s00431-003-1223-x. PMID 12751000.
- ↑ "Stress hyperglycemia, insulin treatment, and innate immune cells". International Journal of Endocrinology 2014: 486403. 8 May 2014. doi:10.1155/2014/486403. PMID 24899891.
- ↑ "The diagnostic value of absolute neutrophil count, band count and morphologic changes of neutrophils in predicting bacterial infections". Medical Principles and Practice 16 (5): 344–347. 2007. doi:10.1159/000104806. PMID 17709921.
- ↑ "Blood cell counts and their correlation with creatine kinase and C-reactive protein in patients with acute myocardial infarction". International Journal of Clinical and Experimental Medicine 5 (1): 50–55. 2012. PMID 22328948.
- ↑ "Absolute neutrophil counts and fibrinogen levels as an aid in the early diagnosis of acute myocardial infarction". Acta Cardiologica 59 (2): 135–140. April 2004. doi:10.2143/ac.59.2.2005167. PMID 15139653.
- ↑ Grune, Jana; Lewis, Andrew J. M.; Yamazoe, Masahiro; Hulsmans, Maarten; Rohde, David; Xiao, Ling; Zhang, Shuang; Ott, Christiane et al. (2022-07-11). "Neutrophils incite and macrophages avert electrical storm after myocardial infarction" (in en). Nature Cardiovascular Research 1 (7): 649–664. doi:10.1038/s44161-022-00094-w. ISSN 2731-0590. PMID 36034743.
- ↑ "Diagnosis of myocardial infarction at autopsy: AECVP reappraisal in the light of the current clinical classification". Virchows Archiv 476 (2): 179–194. February 2020. doi:10.1007/s00428-019-02662-1. PMID 31522288.
"This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/ )" - ↑ "Targeting neutrophils in ischemic stroke: translational insights from experimental studies". Journal of Cerebral Blood Flow and Metabolism 35 (6): 888–901. June 2015. doi:10.1038/jcbfm.2015.45. PMID 25806703.
- ↑ "Genotyping of human neutrophil antigens (HNA) from whole genome sequencing data". BMC Medical Genomics 6 (1): 31. September 2013. doi:10.1186/1755-8794-6-31. PMID 24028078.
- ↑ 63.0 63.1 Ignatov DY (2012). Functional heterogeneity of human neutrophils and their role in peripheral blood leukocyte quantity regulation (PhD). Donetsk National Medical University. doi:10.13140/RG.2.2.35542.34884.
- ↑ "[Functional heterogenicity of human blood neutrophils: generation of oxygen active species"]. Tsitologiia 43 (5): 432–436. 2001. PMID 11517658. https://www.researchgate.net/publication/11827349.
- ↑ "[Neutrophil activation in vitro"]. Tsitologiia 46 (2): 155–158. 2004. PMID 15174354. https://www.researchgate.net/publication/8533160.
- ↑ "[Nitroblue tetrazolium reduction by human blood neutrophils. I. The influence of pH"]. Tsitologiia 47 (6): 549–553. 2005. PMID 16708848. https://www.researchgate.net/publication/7072581.
- ↑ "[Nitroblue tetrazolium reduction by human blood neutrophils. II. The influence of sodium and potassium ions"]. Tsitologiia 47 (6): 554–558. 2005. PMID 16708849. https://www.researchgate.net/publication/7072582.
- ↑ "Single-Cell Transcriptomics of Human and Mouse Lung Cancers Reveals Conserved Myeloid Populations across Individuals and Species". Immunity 50 (5): 1317–1334.e10. May 2019. doi:10.1016/j.immuni.2019.03.009. PMID 30979687.
- ↑ "ITAM signaling by Vav family Rho guanine nucleotide exchange factors regulates interstitial transit rates of neutrophils in vivo". PLOS ONE 4 (2): e4652. 2009. doi:10.1371/journal.pone.0004652. PMID 19247495. Bibcode: 2009PLoSO...4.4652G.
External links
- Neutropenia Information
- Absolute Neutrophil Count Calculator
- Neutrophil Trace Element Content and Distribution
 |  |

