Biology:Cyclin-dependent kinase
This article may incorporate text from a large language model. (October 2025) (Learn how and when to remove this template message) |
| Cyclin-dependent kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 2.7.11.22 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Cyclin-dependent kinases (CDKs) are a predominant group of serine/threonine protein kinases involved in the regulation of the cell cycle and its progression, ensuring the integrity and functionality of cellular machinery. These regulatory enzymes play a crucial role in the regulation of eukaryotic cell cycle and transcription, as well as DNA repair, metabolism, and epigenetic regulation, in response to several extracellular and intracellular signals.[1][2] They are present in all known eukaryotes, and their regulatory function in the cell cycle has been evolutionarily conserved.[3][4] The catalytic activities of CDKs are regulated by interactions with CDK inhibitors (CKIs) and regulatory subunits known as cyclins. Cyclins have no enzymatic activity themselves, but they become active once they bind to CDKs. Without cyclin, CDK is less active than in the cyclin-CDK heterodimer complex.[5][6] CDKs phosphorylate proteins on serine (S) or threonine (T) residues. The specificity of CDKs for their substrates is defined by the S/T-P-X-K/R sequence, where S/T is the phosphorylation site, P is proline, X is any amino acid, and the sequence ends with lysine (K) or arginine (R). This motif ensures CDKs accurately target and modify proteins, crucial for regulating cell cycle and other functions.[7] Deregulation of the CDK activity is linked to various pathologies, including cancer, neurodegenerative diseases, and stroke.[6]
Evolutionary history
CDKs were initially identified through studies in model organisms such as yeasts and frogs, underscoring their pivotal role in cell cycle progression. These enzymes operate by forming complexes with cyclins, whose levels fluctuate throughout the cell cycle, thereby ensuring timely cell cycle transitions. Over the years, the understanding of CDKs has expanded beyond cell division to include roles in gene transcription integration of cellular signals.[7][8]
The evolutionary journey of CDKs has led to a diverse family with specific members dedicated to cell cycle phases or transcriptional control. For instance, budding yeast expresses six distinct CDKs, with some binding multiple cyclins for cell cycle control and others binding with a single cyclin for transcription regulation. In humans, the expansion to 20 CDKs and 29 cyclins illustrates their complex regulatory roles. Key CDKs such as CDK1 are indispensable for cell cycle control, while others like CDK2 and CDK3 are not. Moreover, transcriptional CDKs, such as CDK7 in humans, play crucial roles in initiating transcription by phosphorylating RNA polymerase II (RNAPII), indicating the intricate link between cell cycle regulation and transcriptional management. This evolutionary expansion from simple regulators to multifunctional enzymes underscores the critical importance of CDKs in the complex regulatory networks of eukaryotic cells.[7]
| Species | Name | Original name | Size (amino acids) | Function |
|---|---|---|---|---|
| Saccharomyces cerevisiae | CDK1 | Cdc28 | 298 | All cell-cycle stages |
| Schizosaccharomyces pombe | CDK1 | Cdc2 | 297 | All cell-cycle stages |
| Drosophila melanogaster | CDK1 | Cdc2 | 297 | M |
| CDK2 | Cdc2c | 314 | G1/S, S, possibly M | |
| CDK4 | Cdk4/6 | 317 | G1, promotes growth | |
| Xenopus laevis | CDK1 | Cdc2 | 301 | M |
| CDK2 | 297 | S, possibly M | ||
| Homo sapiens | CDK1 | Cdc2 | 297 | M |
| CDK2 | 298 | G1, S, possibly M | ||
| CDK4 | 301 | G1 | ||
| CDK6 | 326 | G1 |
Notable people
In 2001, the scientists Leland H. Hartwell, Tim Hunt and Sir Paul M. Nurse were awarded the Nobel Prize in Physiology or Medicine for their discovery of key regulators of the cell cycle.[9]
- Leland H. Hartwell (b. 1939): Through studies of yeast in 1971, Heartwell identified crucial genes for cell division, outlining the cell cycle's stages and essential checkpoints to prevent cancerous cell division.[9][10]
- Tim Hunt (b. 1943): Through studies of sea urchins in the 1980s, Hunt discovered the role of cyclins in the regulation of cell cycle phases through their cyclical synthesis and degradation.[9][11]
- Sir Paul M. Nurse (b. 1949): In the mid-1970s, Nurse's studies uncovered the cdc2 gene in fission yeast, which is crucial for the progression of the cell cycle from G1 to S phase and from G2 to M phase. In 1987, he identified the corresponding gene in humans, CDK1, highlighting the conservation of cell cycle control mechanisms across species.[9][12]
CDKs and cyclins in the cell cycle
CDK is one of the estimated 800 human protein kinases. CDKs have low molecular weight, and they are known to be inactive by themselves. They are characterized by their dependency on the regulatory subunit, cyclin. The activation of CDKs also requires post-translational modifications involving phosphorylation reactions. This phosphorylation typically occurs on a specific threonine residue, leading to a conformational change in the CDK that enhances its kinase activity.[13] The activation forms a cyclin-CDK complex which phosphorylates specific regulatory proteins that are required to initiate steps in the cell-cycle.[5]

In human cells, the CDK family comprises 20 different members that play a crucial role in the regulation of the cell cycle and transcription. These are usually separated into cell-cycle CDKs, which regulate cell-cycle transitions and cell division, and transcriptional CDKs, which mediate gene transcription. CDK1, CDK2, CDK3, CDK4 and CDK6 are directly related to the regulation of cell-cycle events, while CDK7 – 13 are associated with transcriptional regulation.[1] Different cyclin-CDK complexes regulate different phases of the cell cycle, known as G0/G1, S, G2, and M phases, featuring several checkpoints to maintain genomic stability and ensure accurate DNA replication.[1][5] Cyclin-CDK complexes of earlier cell-cycle phase help activate cyclin-CDK complexes in later phase.[4]
| CDK | Cyclin partner | Established functions |
|---|---|---|
| CDK1 | cyclin B | M phase transition |
| CDK2 | cyclin A | S/G2 transition |
| CDK2 | cyclin E | G1/S transition |
| CDK3 | cyclin C | G0/G1 and G1/S transitions |
| CDK4, CDK6 | cyclin D | G1/S transition. Phosphorylation of retinoblastoma gene product (Rb) |
| CDK7 | cyclin H | CAK and RNAPII transcription |
CDK structure and activation
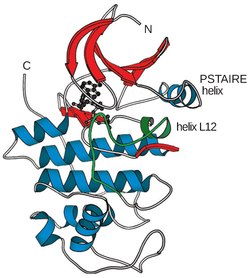
Cyclin-dependent kinases (CDKs) mainly consist of a two-lobed configuration, which is characteristic of all kinases in general. CDKs have specific features in their structure that play a major role in their function and regulation.[2]
- N-terminal lobe (N-lobe): In this part, the inhibitory element known as the glycine-rich G-loop is located. The inhibitory element is found within the beta-sheets in this N-terminal lobe.[4][2] Additionally, there is a helix known as the C-helix. This helix contains the PSTAIRE sequence in CDK1. This region plays a crucial role in regulating the binding between cyclin-dependent kinases (CDKs) and cyclins.[7][2]
- C-terminal lobe (C-lobe): This part contains α-helices and the activation segment, which extends from the DFG motif (D145 in CDK2) to the APE motif (E172 in CDK2). This segment also includes a phosphorylation-sensitive residue (T160 in CDK2) in the so-called T-loop. The activation segment in the C-lobe serves as a platform for the binding of the phospho-acceptor Ser/Thr region of substrates.[7][4][2]
Cyclin binding
The active site, or ATP-binding site, in all kinases is a cleft located between a smaller amino-terminal lobe and a larger carboxy-terminal lobe. Research on the structure of human CDK2 has shown that CDKs have a specially adapted ATP-binding site that can be regulated through the binding of cyclin. Phosphorylation by CDK-activating kinase (CAK) at Thr160 in the T-loop helps to increase the complex's activity. Without cyclin, a flexible loop known as the activation loop or T-loop blocks the cleft, and the positioning of several key amino acids is not optimal for ATP binding.[2][14] With cyclin, two alpha helices change position to enable ATP binding. One of them, the L12 helix located just before the T-loop in the primary sequence, is transformed into a beta strand and helps to reorganize the T-loop so that it no longer blocks the active site. The other alpha helix, known as the PSTAIRE helix, is reorganized and helps to change the position of the key amino acids in the active site.[6][14]
There's considerable specificity in which cyclin binds to CDK. Furthermore, the cyclin binding determines the specificity of the cyclin-CDK complex for certain substrates, highlighting the importance of distinct activation pathways that confer cyclin-binding specificity on CDK1. This illustrates the complexity and fine-tuning in the regulation of the cell cycle through selective binding and activation of CDKs by their respective cyclins.[15][16]
Cyclins can directly bind the substrate or localize the CDK to a subcellular area where the substrate is found. The RXL-binding site was crucial in revealing how CDKs selectively enhance activity toward specific substrates by facilitating substrate docking.[17] Substrate specificity of S cyclins is imparted by the hydrophobic patch, which has affinity for substrate proteins that contain a hydrophobic RXL (or Cy) motif.[4] Cyclin B1 and B2 can localize CDK1 to the nucleus and the Golgi, respectively, through a localization sequence outside the CDK-binding region.[4][16]
Phosphorylation
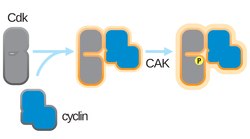
To achieve full kinase activity, an activating phosphorylation on a threonine adjacent to the CDK's active site is required.[18] The identity of the CDK-activating kinase (CAK) that carries out this phosphorylation varies among different model organisms. The timing of this phosphorylation also varies; in mammalian cells, the activating phosphorylation occurs after cyclin binding, while in yeast cells, it occurs before cyclin binding. CAK activity is not regulated by known cell cycle pathways, and it is the cyclin binding that is the limiting step for CDK activation.[4]
Unlike activating phosphorylation, CDK inhibitory phosphorylation is crucial for cell cycle regulation. Various kinases and phosphatases control their phosphorylation state. For instance, the activity of CDK1 is controlled by the balance between WEE1 kinases, Myt1 kinases, and the phosphorylation of Cdc25c phosphatases. Wee1, a kinase preserved across all eukaryotes, phosphorylates CDK1 at Tyr 15. Myt1 can phosphorylate both the threonine (Thr 14) and the tyrosine (Tyr 15). The dephosphorylation is performed by Cdc25c phosphatases, by removing the phosphate groups from both the threonine and the tyrosine.[1][7] This inhibitory phosphorylation helps preventing cell-cycle progression in response to events like DNA damage. The phosphorylation does not significantly alter the CDK structure, but reduces its affinity to the substrate, thereby inhibiting its activity. For the cell cycle to progress, the inhibitory phosphate groups must be removed by the Cdc25 phosphatases to reactivate the CDKs.[7]
CDK inhibitors
A cyclin-dependent kinase inhibitor (CKI) is a protein that interacts with a cyclin-CDK complex to inhibit kinase activity, often during G1 phase or in response to external signals or DNA damage. In animal cells, two primary CKI families exist: the INK4 family (p16, p15, p18, p19) and the CIP/KIP family (p21, p27, p57). The INK4 family proteins specifically bind to and inhibit CDK4 and CDK6 by D-type cyclins or by CAK, while the CIP/KIP family prevent the activation of CDK-cyclin heterodimers, disrupting both cyclin binding and kinase activity.[6][7] These inhibitors have a KID (kinase inhibitory domain) at the N-terminus, facilitating their attachment to cyclins and CDKs. Their primary function occurs in the nucleus, supported by a C-terminal sequence that enables their nuclear translocation.[2]
In yeast and Drosophila, CKIs are strong inhibitors of S- and M-CDK, but do not inhibit G1/S-CDKs. During G1, high levels of CKIs prevent cell cycle events from occurring out of order, but do not prevent transition through the Start checkpoint, which is initiated through G1/S-CDKs. Once the cell cycle is initiated, phosphorylation by early G1/S-CDKs leads to destruction of CKIs, relieving inhibition on later cell cycle transitions.[4] In mammalian cells, the CKI regulation works differently. Mammalian protein p27 (Dacapo in Drosophila) inhibits G1/S- and S-CDKs but does not inhibit S- and M-CDKs.[2]
Ligand-based inhibition methods involve the use of small molecules or ligands that specifically bind to CDK2, which is a crucial regulator of the cell cycle. The ligands bind to the active site of CDK2, thereby blocking its activity. These inhibitors can either mimic the structure of ATP, competing for the active site and preventing protein phosphorylation needed for cell cycle progression, or bind to allosteric sites, altering the structure of CDK2 to decrease its efficiency.[14]

CDK subunits (CKS)
CDKs are essential for the control and regulation of the cell cycle. They are associated with small regulatory subunits regulatory subunits (CKSs). In mammalian cells, two CKSs are known: CKS1 and CKS2. These proteins are necessary for the proper functioning of CDKs, although their exact functions are not yet fully known. An interaction occurs between CKS1 and the carboxy-terminal lobe of CDKs, where they bind together. This binding increases the affinity of the cyclin-CDK complex for its substrates, especially those with multiple phosphorylation sites, thus contributing the promotion of cell proliferation.[20]
Non-cyclin activators
Viral cyclins
Viruses can encode proteins with sequence homology to cyclins. One much-studied example is K-cyclin (or v-cyclin) from Kaposi sarcoma herpes virus (see Kaposi's sarcoma), which activates CDK6. The vCyclin-CDK6 complex promotes an accelerated transition from G1 to S phase in the cell by phosphorylating pRb and releasing E2F. This leads to the removal of inhibition on Cyclin E–CDK2's enzymatic activity. It is shown that vCyclin contributes to promoting transformation and tumorigenesis, mainly through its effect on p27 pSer10 phosphorylation and cytoplasmic sequestration.[21]
CDK5 activators
Two protein types, p35 and p39, responsible for increasing the activity of CDK5 during neuronal differentiation in postnatal development.[22] p35 and p39 play a crucial role in a unique mechanism for regulating CDK5 activity in neuronal development and network formation. The activation of CDK with these cofactors (p35 and p39) does not require phosphorylation of the activation loop, which is different from the traditional activation of many other kinases. This highlights the importance of activating CDK5 activity, which is critical for proper neuronal development, dendritic spine and synapse formation, as well as in response to epileptic events.[22][23]
RINGO/Speedy
The RINGO/Speedy group of proteins can activate CDKs, although their amino acid sequence is not homologous to the cyclin family. Originally identified in Xenopus, these proteins primarily bind to and activate CDK1 and CDK2, despite lacking homology to cyclins. What is particularly interesting, is that CDKs activated by RINGO/Speedy can phosphorylate different sites than those targeted by cyclin-activated CDKs, indicating a unique mode of action for these non-cyclin CDK activators.[24]
Medical significance
CDKs and cancer
The dysregulation of CDKs and cyclins disrupts the cell cycle coordination, which makes them involved in the pathogenesis of several diseases, mainly cancers. Thus, studies of cyclins and cyclin-dependent kinases (CDK) are essential for advancing the understanding of cancer characteristics.[2][25] Research has shown that alterations in cyclins, CDKs, and CDK inhibitors (CKIs) are common in most cancers, involving chromosomal translocations, point mutations, insertions, deletions, gene overexpression, frame-shift mutations, missense mutations, or splicing errors.[2]
The dysregulation of the CDK4/6-RB pathway is a common feature in many cancers, often resulting from various mechanisms that inactivate the cyclin D-CDK4/6 complex. Several signals can lead to overexpression of cyclin D and enhance CDK4/6 activity, contributing toward tumorigenesis.[1][2] Additionally, the CDK4/6-RB pathway interacts with the p53 signaling pathway via p21CIP1 transcription, which can inhibit both cyclin D-CDK4/6 and cyclin E-CDK2 complexes. Mutations in p53 can deactivate the G1 checkpoint, further promoting uncontrolled proliferation.[1][2]
CDK inhibitors and therapeutic potential
Due to their central role in regulating cell cycle progression and cell proliferation, CDKs are considered ideal therapeutic targets for cancer.[25] The following CDK4/6 inhibitors mark a significant advancement in cancer treatment, offering targeted therapies that are effective and have a manageable side effect profile.
- Palbociclib, one of the first CDK4/6 inhibitors approved by the FDA, has become essential in the treatment of HR+/HER2- advanced or metastatic breast cancer, often in combination with endocrine therapy.[26]
- Ribociclib, demonstrating similar efficacy to palbociclib, is also approved for HR+/HER2- advanced breast cancer and offers benefits for a younger patient demographic.[27]
- Abemaciclib stands out by being usable as monotherapy, in addition to combination treatment, for certain HR+/HER2- breast cancer patients. It has also shown effectiveness in treating patients with brain metastases.[27]
- Trilaciclib has proven its value by improving patients' quality of life during cancer treatment by reducing the risk of chemotherapy-induced myelosuppression, a common side effect that can lead to treatment delays and dose reductions.[27]
| Drug | CDKs Inhibited | Condition or disease |
|---|---|---|
| Flavopiridol (alvocidib) | 1, 2, 4, 6, 9 | Acute Myeloid Leukemia (AML) |
| Roscovitine (Seliciclib) | 2, 7, 9 | Pituitary Cushing Disease
Cystic Fibrosis, Advanced Solid Tumors Lung Cancer |
| Dinaclib | 1, 2, 5, 9 | Chronic Lymphocytic Leukemia (CLL)
Breast and Lung Cancers |
| Milciclib | 1, 2, 4, 7 | Hepatocellular Carcinoma (HCC)
Thymic Carcinoma |
| Palbociclib | 4, 6 | Breast Cancer
Head and Neck, Brain, Colon, and other Solid Cancers |
| Ribociclib | 4, 6 | HR+/HER2- Breast Cancer
Prostate, and other Solid Cancers |
| Abemaciclib | 4, 6 | HR+/HER2- Breast Cancer
Lung, Brain, Colon, and other Solid Cancers |
| Meriolin | 1, 2, 5, 9 | Neuroblastoma, Glioma, Myeloma, Colon Cancer |
| Variolin B | 1, 2, 5, 9 | Murine Leukemia |
| Roniciclib | 1, 2, 4, 7, 9 | Lung and Advanced Solid Cancers |
| Meridianin E | 1, 5, 9 | Larynx Carcinoma
Myeloid Leukemia |
| Nortopsentins | 1 | Malignant Pleural Mesothelioma (MPM) |
Challenges and future potential
Complications of developing a CDK drug include the fact that many CDKs are not involved in the cell cycle, but other processes such as transcription, neural physiology, and glucose homeostasis.[30] More research is required, however, because disruption of the CDK-mediated pathway has potentially serious consequences; while CDK inhibitors seem promising, it has to be determined how side-effects can be limited so that only target cells are affected. As such diseases are currently treated with glucocorticoids.[31] The comparison with glucocorticoids serves to illustrate the potential benefits of CDK inhibitors, assuming their side effects can be more narrowly minimized or targeted.[32]
See also
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 "The Roles of Cyclin-Dependent Kinases in Cell-Cycle Progression and Therapeutic Strategies in Human Breast Cancer". International Journal of Molecular Sciences 21 (6): 1960. March 2020. doi:10.3390/ijms21061960. PMID 32183020.
- ↑ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 2.11 "The driving power of the cell cycle: cyclin-dependent kinases, cyclins and their inhibitors". Bratislavske Lekarske Listy 124 (4): 261–266. 2023. doi:10.4149/BLL_2023_039. PMID 36598318.
- ↑ "Genome-wide identification, evolutionary and expression analysis of the cyclin-dependent kinase gene family in peanut". BMC Plant Biology 23 (1). January 2023. doi:10.1186/s12870-023-04045-w. PMID 36658501. Bibcode: 2023BMCPB..23...43S.
- ↑ 4.0 4.1 4.2 4.3 4.4 4.5 4.6 4.7 4.8 The Cell Cycle: Principles of Control. London: New Science Press Ltd. 2007. pp. 2–54, 196–266. ISBN 978-0-9539181-2-6.
- ↑ 5.0 5.1 5.2 5.3 Essential Cell Biology (5th ed.). W. W. Norton & Company. 2019. pp. 613–627. ISBN 9780393679533.
- ↑ 6.0 6.1 6.2 6.3 6.4 "Cyclin-Dependent Kinases (CDK) and Their Role in Diseases Development-Review". International Journal of Molecular Sciences 22 (6): 2935. March 2021. doi:10.3390/ijms22062935. PMID 33805800.
- ↑ 7.0 7.1 7.2 7.3 7.4 7.5 7.6 7.7 "Cyclin-dependent kinases". Genome Biology 15 (6). June 30, 2014. doi:10.1186/gb4184. PMID 25180339.
- ↑ "Cyclin/Forkhead-mediated coordination of cyclin waves: an autonomous oscillator rationalizing the quantitative model of Cdk control for budding yeast". npj Systems Biology and Applications 7 (1). December 2021. doi:10.1038/s41540-021-00201-w. PMID 34903735.
- ↑ 9.0 9.1 9.2 9.3 "A Journey through Time on the Discovery of Cell Cycle Regulation". Cells 11 (4): 704. February 2022. doi:10.3390/cells11040704. PMID 35203358.
- ↑ "The Nobel Prize in Physiology or Medicine 2001" (in en-US). https://www.nobelprize.org/prizes/medicine/2001/hartwell/facts/.
- ↑ "The Nobel Prize in Physiology or Medicine 2001" (in en-US). https://www.nobelprize.org/prizes/medicine/2001/hunt/facts/.
- ↑ "The Nobel Prize in Physiology or Medicine 2001" (in en-US). https://www.nobelprize.org/prizes/medicine/2001/nurse/facts/.
- ↑ "Identifying in vivo targets of cyclin-dependent kinase inhibitors by affinity chromatography". Biochemical Pharmacology. Cell Signaling, Transcription and Translation as Therapeutic Targets 64 (5–6): 819–825. September 2002. doi:10.1016/S0006-2952(02)01144-9. PMID 12213575.
- ↑ 14.0 14.1 14.2 "Insights on Structural Characteristics and Ligand Binding Mechanisms of CDK2". International Journal of Molecular Sciences 16 (5): 9314–9340. April 2015. doi:10.3390/ijms16059314. PMID 25918937.
- ↑ "Distinct activation pathways confer cyclin-binding specificity on Cdk1 and Cdk2 in human cells". Molecular Cell 32 (5): 662–672. December 2008. doi:10.1016/j.molcel.2008.10.022. PMID 19061641.
- ↑ 16.0 16.1 "The Cyclin-dependent kinase 1: more than a cell cycle regulator". British Journal of Cancer 129 (11): 1707–1716. November 2023. doi:10.1038/s41416-023-02468-8. PMID 37898722.
- ↑ "Structural insights into the functional diversity of the CDK-cyclin family". Open Biology 8 (9). September 2018. doi:10.1098/rsob.180112. PMID 30185601.
- ↑ "Cyclins and cyclin-dependent kinases: from biology to tumorigenesis and therapeutic opportunities". Journal of Cancer Research and Clinical Oncology 149 (4): 1585–1606. April 2023. doi:10.1007/s00432-022-04135-6. PMID 35781526.
- ↑ "Identification of selective cyclin-dependent kinase 2 inhibitor from the library of pyrrolone-fused benzosuberene compounds: an in silico exploration". Journal of Biomolecular Structure & Dynamics 40 (17): 7693–7701. October 2022. doi:10.1080/07391102.2021.1900918. PMID 33749525. https://figshare.com/articles/journal_contribution/Identification_of_selective_cyclin-dependent_kinase_2_inhibitor_from_the_library_of_pyrrolone-fused_benzosuberene_compounds_an_in_silico_exploration/14259911/1/files/26943209.pdf.
- ↑ "Cyclin-dependent kinase regulatory subunit 1 promotes cell proliferation by insulin regulation". Cell Cycle 14 (19): 3045–3057. July 22, 2015. doi:10.1080/15384101.2015.1053664. PMID 26199131.
- ↑ "Viral cyclin promotes KSHV-induced cellular transformation and tumorigenesis by overriding contact inhibition". Cell Cycle 13 (5): 845–858. March 1, 2014. doi:10.4161/cc.27758. PMID 24419204.
- ↑ 22.0 22.1 "p39 Is Responsible for Increasing Cdk5 Activity during Postnatal Neuron Differentiation and Governs Neuronal Network Formation and Epileptic Responses". The Journal of Neuroscience 36 (44): 11283–11294. November 2016. doi:10.1523/JNEUROSCI.1155-16.2016. PMID 27807169.
- ↑ "Cdk5 activation promotes Cos-7 cells transition towards neuronal-like cells". Translational Neuroscience 14 (1). January 2023. doi:10.1515/tnsci-2022-0318. PMID 37901140.
- ↑ "RINGO/Speedy proteins, a family of non-canonical activators of CDK1 and CDK2". Seminars in Cell & Developmental Biology. 1. Cyclins edited by Josep Clotet 107: 21–27. November 2020. doi:10.1016/j.semcdb.2020.03.010. PMID 32317145.
- ↑ 25.0 25.1 "A review on the role of cyclin dependent kinases in cancers". Cancer Cell International 22 (1). October 2022. doi:10.1186/s12935-022-02747-z. PMID 36266723.
- ↑ "Coming of Age: Targeting Cyclin K in Cancers". Cells 12 (16): 2044. August 2023. doi:10.3390/cells12162044. PMID 37626854.
- ↑ 27.0 27.1 27.2 "CDK inhibitors from past to present: A new wave of cancer therapy". Seminars in Cancer Biology 88: 106–122. January 2023. doi:10.1016/j.semcancer.2022.12.006. PMID 36565895.
- ↑ "Inhibitors of Cyclin-Dependent Kinases: Types and Their Mechanism of Action". International Journal of Molecular Sciences 22 (6): 2806. March 2021. doi:10.3390/ijms22062806. PMID 33802080.
- ↑ "Cyclin dependent kinase (CDK) inhibitors as anticancer drugs: Recent advances (2015-2019)". Bioorganic & Medicinal Chemistry Letters 29 (20). October 2019. doi:10.1016/j.bmcl.2019.126637. PMID 31477350.
- ↑ "Fueling the Cycle: CDKs in Carbon and Energy Metabolism". Frontiers in Cell and Developmental Biology 6. August 17, 2018. doi:10.3389/fcell.2018.00093. PMID 30175098.
- ↑ "An Overview of the Safety Profile and Clinical Impact of CDK4/6 Inhibitors in Breast Cancer-A Systematic Review of Randomized Phase II and III Clinical Trials". Biomolecules 13 (9): 1422. September 2023. doi:10.3390/biom13091422. PMID 37759823.
- ↑ "The long winding road to the safer glucocorticoid receptor (GR) targeting therapies". Oncotarget 13: 408–424. February 18, 2022. doi:10.18632/oncotarget.28191. PMID 35198100.
External links
- Cyclin-Dependent+Kinases at the US National Library of Medicine Medical Subject Headings (MeSH)
 |  |
